КО-диеты - наиболее раннее и многосторонне изученное направление. Суть постановки опыта заключается в том, что, начиная с определенного возраста, количество потребляемой пищи принудительно снижают (обычно на 20-60 %) Если подобное снижение достаточно «жестко» и начато с ранних постнатальных периодов, то время от времени животных переводят на нормальный рацион с тем, чтобы завершился их рост. Поскольку при КО-диетах пропорционально снижается поступление в организм как определяющих калорийность продуктов (в основном углеводы, жиры), так и множества других биологически важных соединений, то почти всегда к КО-диетам добавляют недостающие количества наиболее необходимых веществ (витамины, микроэлементы). В некоторых случаях компенсируют и ряд других составляющих рациона, например белки, и нередко называют подобную диету качественно полноценной, но калорийно недостаточной (Никитин, 1984) Учитывая сложность, многокомпонентность пищевых продуктов, вряд ли подобную компенсацию можно считать полноценной.
Первые опыты с КО-диетами начинали на животных с возможно ранних этапов постнатального онтогенеза, и основным критерием эффективности различных диет считали степень задержки роста Однако в 40-е годы эти представления были поставлены под сомнения и их ошибочность была экспериментально доказана Карлсоном из Чикагского университета, который показал, что увеличение ПЖ с помощью КО-диет может быть достигнуто и при их применении после завершения роста (цит. по: Ross, 1978). Значимость эгого открытия трудно переоценить для практического применения КО-диет. Дело в том, что эффективное ограничение калорий у экспериментальных животных вряд ли может быть применено у человека с раннего детства, так как гораздо меньшие снижения калорий у детей вызывают серьезные нарушения ВНД, повышают чувствительность к болезням и т. д. (Ross, 1978). Продление жизни КО-диетами, начатыми в зрелом возрасте, конечно, более перспективно для клинического внедрения.
Как показали исследования ряда авторов, пролонгирующий жизнь эффект КО-диет зависит от начального возраста применения и прогрессивно падает по мере увеличения возраста. Например, в исследованиях Гудрика и соавт. (Goodrick et al., 1982a), проведенных на долгоживущей (C57BL/6J) и краткоживущей (A/J) линиях мышей было показано, что примерно двухкратное снижение калорийности диеты, начатое с 45-, 180- и 300-дневного возраста, увеличивает СПЖ соответственно на 27, 11, 0 % у мышей линии C57BL/6J, у A/J - на 12 и 2 %, сокращает на 14 %. Об отрицательном влиянии КО-диет на ПЖ крыс с «позднего» (365-суточного) возраста свидетельствуют данные Барроуза и Роедера (Barrows, Roeder, 1965), в опытах которых СПЖ снижалась на 8 %. Однако не всегда КО-диеты, начатые в зрелом возрасте, оказывались малоэффективными. Например, в исследованиях Стухликовой и соавт. (Stuchlikova et al., 1975) снижение калорийности питания путем перевода подопытных животных на режим кормления через день обеспечивало рост СПЖ мышей, крыс и золотистых хомячков примерно на 17, 32 и 9 % при применении КО-диеты с молодого возраста и на 29, 43, 30 % при переводе на КО-диету в годовалом возрасте. Некоторые авторы полагают, что для взрослых животных с их сниженными адаптационно-регуляторными возможностями используемые режимы ограничения диеты, оптимальные для молодых, являются слишком большим испытанием и зачастую губительны. Поэтому при работе на животных старших возрастных групп необходимо более умеренное и постепенное ограничение диеты. Эффективность подобного подхода была продемонстрирована в работе Вейндрача и Волфорда (Weindruch, Walford, 1982), которые двухступенчатым снижением калорийности пищи примерно на 45 %, начиная с 400-суточного возраста, увеличили СПЖ и МПЖ долгоживущих мышей линии C57BL/6J на 20 и 25 %, а у «сверхдолгоживущих» мышей C57BL/10 SnxC3H/HeDiSn F1-Ha 12 и 11 % соответственно.
Довольно неожиданными оказались результаты опытов с КО-диетами в несколько иной постановке опытов: снижение калорийности на первой половине и неограниченное питание во второй половине жизни. Так, в опытах Стухликовой и соавт. (Stuchlikova et al., 1975) мыши, крысы и хомячки в подопытных группах пережили своих контрольных сородичей на 39, 61 и 50 %. В аналогичной серии опытов на самцах Вистар Боучен и соавт. (Beauchane et al., 1979) наблюдали увеличение СПЖ на 13 %. После перехода в режим неограниченного кормления животные переедали и быстро жирели. Тем не менее по ПЖ они не уступали и даже несколько превосходили не только контрольных, но и животных, получавших КО-диету в течение всей жизни. Как полагает Станкард (Stunkard, 1976), если результаты этих опытов подтвердятся, то придется допустить, что не все виды ожирения отрицательно влияют на ПЖ и взаимоотношения между этими параметрами не так однозначны, как это представлялось до недавнего времени.
Особый интерес представляют комплексные исследования, когда в одинаковых условиях изучается влияние разных вариантов КО- и БО-диет не только на ПЖ, но и на возрастные изменения биологических показателей, возрастную патологию и др. Одна из таких работ с использованием 568 самцов крыс F344 была проведена сотрудниками Техасского университета (Yu et al., 1985; Maeda et al., 1985). Было показано, что ограничение количества пищи на 60 % приводит к увеличению СПЖ на 51 %, а МПЖ - на 38 %, если оно было начато с 6-недельного возраста и соответственно на 34 и 38 %, если ограничение диеты было начато с 6-месячного возраста. Ограничение только в период от 6 нед до 6 мес оказалось малоэффективным (15 и 11 %). Снижение содержания только белка с 21 до 12.6 % (при одинаковом с контрольными крысами калораже) с 6-недельного возраста также малоэффективно (рост СПЖ - 15, МПЖ - 3 %). Безусловным достоинством этой работы является то, что, обратив особое внимание на возрастную динамику массы тела и количества потребляемой пищи, авторы опровергли неверные представления, будто при КО-диетах животные потребляют меньше калорий. Из расчета на единицу тощей массы тела количество потребляемых калорий было одинаковым (примерно 135-145 ккал/сут • кг) при разных постановках опыта. В зависимости от степени и момента ограничения снижалась только масса тела подопытных животных. Интересно, что снижение массы тела не сопровождалось снижением массы большинства внутренних органов - сердца, селезенки, почек, легких и тестикул. Только масса печени коррелировала с массой тела. При КО-диете с возрастом спонтанная локомоторная активность у подопытных крыс в отличие от контрольных существенно не изменялась, а АД в обеих группах изменялось одинаково, прогрессивно возрастая примерно со 100 до 140 мм рт. ст.
Таким образом, из представленных данных следует, что KOдиеты, особенно при их применении до половозрелого возраста, способны значительно увеличивать ПЖ мелких лабораторных грызунов. Вместе с тем далеко не всегда КО-диеты даже оптимальной силы и начала применения приводили к увеличению ПЖ. Наряду с опытами, в которых СПЖ и МПЖ увеличиваются на 60-80 % (Ross, 1959, 1978; Stuchlikova et al., 1975; Goodrick et al., 1982b), встречаются работы, в которых нет данных, указывающих на сколько-нибудь заметное удлинение жизни. Особенно незначительным оказывается эффект в опытах с долгоживущими линиями или популяциями животных. Так, в 4 хорошо продуманных опытах на мышах C57BL/6J Чени и соавт. (Cheney et al., 1980), начиная примерно 40 %-ное ограничение диеты с 21-24-дневного возраста, не наблюдали существенного роста СПЖ.
Для опытов с пролонгированием жизни с помощью КО-диет примерно в 60 % случаев характерно более существенное увеличение МПЖ, чем СПЖ. В этом плане КО-диеты выгодно отли- 9 В В Фролькис, X К Мурадян чаются от других известных геропротекторов, которые обычно на фоне значительного увеличения СПЖ мало влияют на МПЖ. Подобные отличия могут быть объяснены тем, что жесткие ограничения пищи приводят к значительной перестройке адаптационно-регуляторных механизмов, к изменению соотношения процессов старения и витаукта. У ряда животных адаптационные возможности не перестраиваются, и это нередко кончается их гибелью. Однако из-за менее выраженного возрастного роста смертности подопытные животные постепенно превосходят по ПЖ контрольных. Так, по оценкам Сейчер (Sacher, 1977), показатель смертности в начале эксперимента (R0) в группе подопытных крыс был почти в 3 раза выше (8 • 10~6день~ ), чем у контрольных животных (2.7 • 10~6 день" ). Однако если в группе подопытных смертность удваивалась через каждые 105-130 дней, то в контрольной - вдвое быстрее, что, по мнению Сейчер, - наилучшее доказательство «истинно геропротекторного» действия КО-диет. В результате подобных разнонаправленных изменений смертности подопытных животных на первой и второй половинах эксперимента обычно рост СПЖ оказывается меньше, чем МПЖ-
Как указывалось, важно не только «количество», но и «качество» жизни, характер изменения биологического возраста, знание механизмов пролонгирующего действия КО-диеты. Уже по внешнему виду животные, находящиеся на ограниченной диете, отличались от контрольных - они были более подвижны, в возрасте 24 мес у них была мягкая белая шерсть. Морфологический анализ показал, что позже во многих органах возникают деструктивные изменения. У крыс с ограниченной диетой задержаны возрастные изменения белков в крови, в структуре коллагена, изменяется обычное соотношение коллагена и эластина в коже (Никитин, 1984). Существенно отличаются у подопытных животных темп и тип возрастных сдвигов в геноме, в структуре хроматина. Гормональная регуляция при КО-диете изменяется значительно и разнонаправленно, что позволило В. H. Никитину (1984) писать о возникновении при этом новой эндокринной ситуации.
При ограниченной диете растет подвижность животных, их возбудимость, агрессивность, снижается порог судорожной готовности, падает устойчивость к ряду стрессов, отодвигается время полового созревания, в более позднем возрасте сохраняется репродуктивная способность.
Ограниченная диета существенно влияет на развитие возрастной патологии. Так, у КО-животных менее выражена старческая гипертрофия миокарда, значительно снижается частота патологических нарушений в клубочках и канальцах почек, предотвращается старческая протеинурия (Wyndham et al., 1983) и т. д. В настоящее время считается установленным, что КО-диеты задерживают или ослабляют развитие многих заболеваний почек (Everitt et al., 1982), диабета (Gerritsen, 1976), атеросклероза (Hazzard, 1976), нарушения костной ткани (KaIu et al., 1984).
Специально следует остановиться на влиянии КО-диет на частоту и возрастную динамику возникновения и протекания опухолей. Еще в первых исследованиях Маккея было обнаружено, что КО-диеты приводят к снижению частоты опухолей аденогипофиза и легких. Далее было показано, что при КО-диетах, помимо лимфом, снижается частота злокачественных новообразований молочной железы, печени, легких, почек, сердца, простаты и ряда других тканей (Gerbase-DeLima et al., 1975). Чем жестче КО-диета, тем заметнее снижение частоты или задержка возникновения опухолей. Например, по данным Росса (Ross, 1978), у крыс SD, получавших в течение всей жизни КО-диету, смертность от опухолей была в 10 раз ниже, в том числе количество опухолей в сердце падало в 50 раз (!), простате - в 20 раз, легких - в 9 раз и т. д. Однако в большинстве случаев КО-диеты существенно не влияют на частоту, а лишь отодвигают сроки наступления различных патологий и, следовательно, не устраняют, а лишь задерживают проявление причин старения.
Ограниченная диета резко изменяет пищевое поведение животных, функцию пищевого центра, по И. П. Павлову, и в первую очередь функцию ядер гипоталамуса - не только вентрального и латерального, но и переднего отдела гипоталамуса. Доказательством роли нейрогормональных сдвигов является изменение при КО-диете гипоталамо-гипофизарных взаимоотношений: усиление инкреции кортикотропина, снижение тиреотропина, стабильный до старости уровень фолликулостимулирующего и лютеинизирующего гормона в крови. Это приводит к последующим изменениям секреторной активности других желез внутренней секреции. Так, по данным В. H. Никитина (1962, 1984), у подопытных животных масса надпочечников, ширина клубочковой зоны в 2 раза больше, чем у контрольных. У подопытных крыс усилена секреция кортикостероидов и почти на 40 % выше способность синтеза их надпочечниками. Наряду с этим в течение всей жизни снижена концентрация гормонов щитовидной железы, инсулина, тестостерона. О ведущей роли гипоталамуса в механизме наступающих при КО-диете сдвигов свидетельствуют изменения в репродуктивной способности животных. По данным И. А. Аршавского (1979), у контрольных крыс половое созревание начинается не позднее 55-60-го дня жизни, у подопытных - через 6-7 мес. Под влиянием КО-диеты семенники значительно отстают в своем развитии. Однако в старости в них менее выражены деструктивные сдвиги, более длительно сохраняется репродуктивная способность.
По нашим данным, существенное значение при старении приобретает феномен так называемой «гипоталамической дезинформации». Оказалось, что с возрастом изменяется чувствительность гипоталамуса к гормонам - к одним растет, к другим падает, к третьим не изменяется. Все это приводит к неверной информации центров о состоянии внутренней среды организма, к нарушениям ее регуляции. Чрезвычайно важно, как показал В. H. Никитин (1984), что при КО-диете длительно сохраняется реактивность неироэндокриннои системы, способность реагировать на адекватные раздражения. Так, у контрольных животных реакция гипофиза на кортиколиберин резко падает к 12 мес, а у подопытных практически не изменяется. Высоким сохраняется и ингибирующее влияние (отрицательная обратная связь) гидрокортизона на систему гипоталамус-гипофиз-кора надпочечников.
В феномене увеличения ПЖ под влиянием КО-диеты имеет значение и другой уровень регуляции - генорегуляторный. Это доказывается прямыми данными об изменении системы, обеспечивающей регуляцию и считывание генетической информации - хроматина (Никитин, 1984). При КО-диете «сдержано» увеличение содержания гистонов, к старости почти вдвое нарастает содержание негистоновых белков, изменяется их фосфорилирование, ацетилирование, метилирование.
Менее резкие изменения наступают в активности нейтральной и кислой ДНКазы, снижается активность РНК-полимераз А и Б, менее выражено обогащение хроматина фосфолипидами и др. (Никитин, 1984; Клименко, 1984). Весь этот комплекс сдвигов сказывается на реализации генетической информации и в связи с этим на темпе роста, ПЖ животных.
Итак, пролонгирующий эффект ограниченных диет связан с изменением процессов регуляции на разных уровнях биологической организации. Чем стабильнее эти механизмы регуляции, чем они более косны, тем труднее перевести их на новый уровень и вызвать пролонгирующий эффект. КО-диеты изменяют взаимоотношения процессов старения и витаукта. Более медленный темп возрастного развития приводит к тому, что успевают отмобилизоваться процессы витаукта, которые более выраженно превалируют над старением в отдельные возрастные периоды.
У теплокровных животных КО-диеты считаются если не единственным, то по крайней мере самым эффективным способом продления жизни. Наверно поэтому разработке путей продления жизни именно на этой модели было уделено наибольшее внимание. И, как это характерно для интенсивно развивающихся областей науки, критический анализ полученного экспериментального материала и тем более его последующее использование в логическом синтезе значительно отстает от потока экспериментальных работ.
Вот почему, не претендуя на полноту и всесторонность анализа, мы все же решились на обобщение некоторых положений, которые следуют из совместной оценки результатов разных авторов. При этом мы понимаем «механистичность» подобного подхода, возможность упущения каких-то важных деталей, «нивелировки нюансов» эксперимента. Но именно в этом одновременно видим и преимущество - концентрацию внимания на основной задаче этих экспериментов
Прежде всего обращает на себя внимание экспоненциальный рост числа оригинальных исследований. Из всех известных нам 78 серий опытов по изучению влияния КО-диет на ПЖ млекопитающих 10 были проведены до 1970 г , 27 - в 70-х годах и 41 - только за первую половину 80-х годов, т. е. число серий опытов по изучению ПЖ более чем удваивается за каждое десятилетие (рис. 22). В последние годы резко вырос разброс результатов разных авторов. Если в работах до 80-х годов пролонгирующий эффект КО-диет обнаруживали без исключения авторы всех работ и предметом дискуссии мог быть только количественный анализ эффектов, то в исследованиях 80-х годов в целом ряде случаев не был обнаружен сколько-нибудь существенный пролонгирующий эффект, а некоторые авторы, наоборот, обнаруживали рекордно большие увеличения ПЖ. В результате этого коэффициент вариации процентного роста СПЖ в 80-х годах увеличился в 4 раза, а МПЖ - в 6 раз по сравнению с более ранними исследованиями (рис. 22).

Рис. 22. Влияние КО-диет на изменения СПЖ и МПЖ, а также их коэффициентов вариации.
Работы проведены / - до 1970 г , 2 - с 1970 по 1979 г , 3 - с 1980 по 1984 г.
Рис. 23. Гистограмма распределения прироста СПЖ в опытах с КО-диетами.
Подобное резкое увеличение коэффициента вариации свидетельствует о вступлении исследований КО-диет в критическую фазу, и только после ее прохождения, очевидно, может быть проведена объективная оценка истинных возможностей этого класса геропротекторов. В доступном нам массиве работ были изучены СПЖ и МПЖ 195 популяций животных, причем исследования на самцах проводились примерно в 3 раза чаще, у них и пролонгирующий эффект был примерно в 3 раза выше и в 2 раза стабильнее (судя по коэффициенту вариации), чем у самок. В среднем под влиянием КО-диеты СПЖ увеличивалась на (22±3) % с размахом от -34 до +83 %. Примерно такие же данные наблюдаются для МПЖ, которая в среднем увеличивалась на (23±6) % с размахом от -42 до +83 %. Судя по гистограмме, форма распределения роста ПЖ под влиянием КО-диет близка к нормальному или пуассоновскому распределению, т. е. пролонгирующий эффект диеты, очевидно, определяется влиянием множества переменных стохастической природы (рис. 23). Из этого же рисунка видно, что примерно в 90 % случаев эффект КО-диет положительный, а модальный прирост СПЖ соответствует 10-20 %. По всей вероятности, эти результаты несколько завышены, так как большинство исследователей не учитывает смертности на первых этапах эксперимента, сразу после применения калорийного ограничения. Между тем это может нарушить правило рандомизации сравниваемых популяций контрольных и подопытных групп: на этом этапе, очевидно, погибает наиболее слабая, маложизнеспособная часть подопытных животных, данные которых в отличие от контрольной группы выпадают из окончательного расчета.
При рассмотрении работ создавалось впечатление, что пролонгирующий эффект КО-диет в значительной степени зависит от возраста, с которого начато воздействие. Из анализа 49 серий опытов, представленного на рис. 24, видно, что наибольший пролонгирующий эффект наблюдался при начале действия КОдиет до достижения половозрелого возраста: для СПЖ - с 24- 25-суточного, для МПЖ - с 21-суточного. Рост этих показателей достигает 30 % для СПЖ и 37 % для МПЖ при коэффициенте вариации, в обоих случаях близким к 1. При применении КОдиет после завершения роста (начиная с возраста более 300 сут) пролонгирующий эффект снижается и, что особенно заметно, резко увеличивается размах (от -34 до +43 %) и коэффициент вариации (в 2-3 раза по сравнению с предыдущими группами).
Как отмечалось (Sacher, 1977), «истинными» геропротекторами, замедляющими темпы старения, можно считать только те факторы, которые увеличивают ПЖ генетически полноценных, долгоживущих популяций вида, а не короткоживущих линий животных. Определенную ясность в эту проблему вносит сопоставление эффекта КО-диеты с СПЖ и МПЖ контрольных групп животных. Результаты подобного анализа, проведенного нами, представлены на рис. 25. Из этих данных отчетливо видно, что по мере увеличения ПЖ контрольных животных пролонгирующий эффект КО-диет (в % или абсолютном выражении) заметно снижается. Причем эта зависимость одинаково хорошо описывается как ниспадающей экспонентой, так и прямой с отрицательным коэффициентом регрессии. В последнем случае, например, зависимость между приростом СПЖ (у, А %) при КО-диетах и СПЖ {х, сут) контрольных животных имеет вид: i/= 102 - OAx. Другими словами, из анализа работ разных авторов следует, что при увеличении СПЖ контрольных животных на каждые 10 дней стимулирующий эффект КО-диет «снижается» на 1 %. Подобная обратная зависимость отчетливо проявляется и для МПЖ. При ограничении калорий, начатом не позднее двухмесячного возраста, если СПЖ контрольных животных не превышала 550 дней, то СПЖ подопытных увеличивалась в среднем на 53%, 550-649 дней - на 33, 650-749 дней - на 23, 750-850 дней - на 16 и более 850 дней - на 7%. Аналогичные величины для МПЖ со 100-дневным интервалом от менее 850 до более 1150 дней составляли 61, 38, 28, 11 и 1 %. В абсолютном же выражении СПЖ и МПЖ этих групп мало отличались, составляя, например, для МПЖ 1290, 1238, 1275, 1225 и 1214 дней. Другими словами, за высоким процентным ростом СПЖ или МПЖ обычно кроются сравнительно одинаковые абсолютные величины ПЖ, и числовое выражение процентного роста чаще определяется отклонениями ПЖ контрольных животных, а не подопытных.
Рис. 24. Зависимость прироста СПЖ и МПЖ в опытах с КО-диетами от возраста, с которого начато воздействие.
Интересна и динамика этих показателей за последние десятилетия. Если, по данным до 1970 г. при КО-диетах СПЖ и МПЖ увеличивались в среднем на 32 и 72 %, то в 70-х годах эти величины снизились до 27 и 21 %, а в первой половине 80-х-до 15 и 16 % соответственно (рис. 22). Подобное снижение пролонгирующего эффекта, возможно, является следствием улучшения условий содержания контрольных животных и увеличения их ПЖ, а также более критического подхода ряда исследователей к решению такой важной задачи, как продление жизни млекопитающих
Уже первые исследования показали, что до определенного предела (до 60-70 %) рост ПЖ тем больше, чем больше степень ограничения рациона. Так, по данным Росса (Ross, 1959), при снижении калорийности диеты на 40 % СПЖ крыс увеличивалась на 14 %, а при ограничении на 60 % рост СПЖ составлял 27 %. Дальнейшее увеличение степени ограничения, например до 75 %, не приводило к увеличению геропротекторного эффекта. Исходя из этих и ряда других данных, закрепилось представление о том, что наибольший пролонгирующий эффект соответствует ограничению диеты на 40-60 %, и в 85 % случаев применялись ограничения калорийности рациона только в этом узком диапазоне. Обработка результатов 26 серий опытов, проведенных на крысах линии SD, со степенью ограничения калорийности рациона до 75 % показала, что зависимость между ростом СПЖ (у, %) и среднесуточной калорийностью диеты (х, ккал/г • сут) достаточно хорошо описывается линейной регрессией' t/= (1226± ±61) - (3 81 ±0 62)* с множественным коэффициентом детерминации Z? = 61 %. Из этого уравнения видно, что более 60 % дисперсии СПЖ укладывается в эту зависимость и коэффициент регрессии (в скобках приведена его величина и стандартное отклонение) значимо отличается от нуля, причем при снижении калорийности рациона на каждую калорию следует ожидать увеличения СПЖ на 3.81 сут.
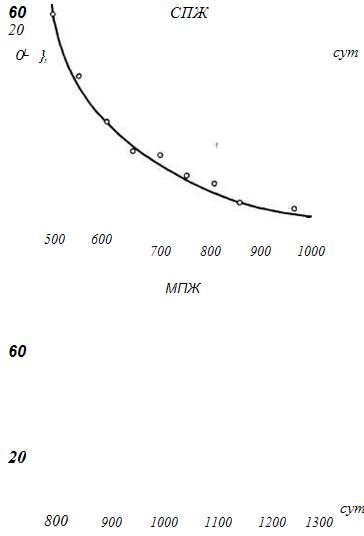
Рис. 25. Зависимость прироста СПЖ и МПЖ от ПЖ интактных животных в опытах с КО-диетами.
Аналогичное уравнение для МПЖ показало, что зависимость между МПЖ и калорийностью диеты еще более отчетливая: у= (21ilO± 114) - (8.75± 1.06)x. В эту зависимость укладывается 92 % дисперсии МПЖ, а сокращение каждой калории должно привести к увеличению МПЖ на 8.75 сут (рис. 26).
Опыты с КО-диетами интересны не только для оценки максимальных возможностей продления жизни млекопитающих. Они могут быть рассмотрены как модельные для изучения и проверки пригодности основных закономерностей старения и путей продления жизни млекопитающих. В частности, на этой модели интересно сопоставление сдвигов СПЖ и МПЖ - факт, на который обращали внимание многие исследователи (Sacher, 1977).
В разных когортах интактных животных, и тем более при различных экспериментальных воздействиях, сдвиги СПЖ и МПЖ, как правило, оказываются однонаправленными, но поразному выраженными, и парный коэффициент корреляции между СПЖ и МПЖ всегда ниже единицы В частности, согласно нашим оценкам, у крыс парный коэффициент корреляции между СПЖ и МПЖ составляет г2д = 0 93 Аналогичные коэффициенты между ростом СПЖ и калорийностью рациона г12 = -0 78, а для МПЖ г13 = -0 95 Судя по величинам г12 и /-J3 оба показателя ПЖ растут по мере снижения калорийности рациона Но эта корреляция может быть формальной, и даже высокие значения коэффициента корреляции еще не означают, что между калорийностью и механизмами СПЖ и МПЖ существует причинно-следственная зависимость Одной из возможностей более корректной оценки причинно-следственных взаимоотношений является замена парных коэффициентов корреляции частными коэффициентами корреляции, что позволяет исключить из корреляционной связи вклад определенной группы факторов.
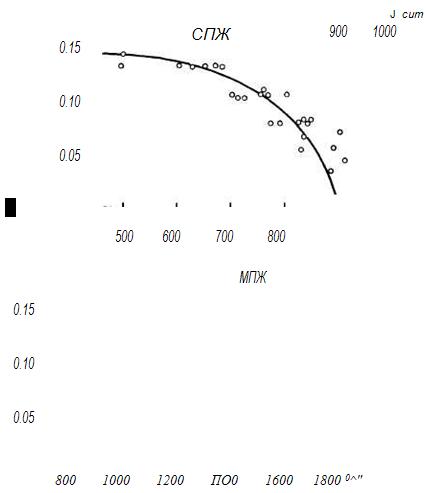
Рис 26. Зависимость СПЖ и МПЖ крыс линии SD от калорийности диеты.
Проведенные нами расчеты показали, что частный коэффициент корреляции между калорийностью и МПЖ (г 13.2) приближается к -1, а аналогичный показатель для СПЖ не только не уменьшается по сравнению с парным коэффициентом корреляции, но и меняет знак на противоположный - r{2. 3 = +0.781 Из представленных данных следует, что «истинная» отрицательная коррелятивная зависимость существует только между калорийностью рациона и МПЖ, а для СПЖ, наоборот, подобная зависимость положительная Следовательно, приведенная выше отрицательная коррелятивная зависимость между калорийностью рациона и ростом СПЖ является формальной и обусловлена лишь более выраженной связью СПЖ с МПЖ, чем СПЖ с калорийностью Нам представляется, что подобный дифференцированный подход при выяснении сдвигов СПЖ и МПЖ может оказаться полезным для оценки влияния и других геропротекторов и моделей ПЖ.
См. также:  Белково-ограниченная диета.
Белково-ограниченная диета.
 Обсудить на форуме
Обсудить на форуме

