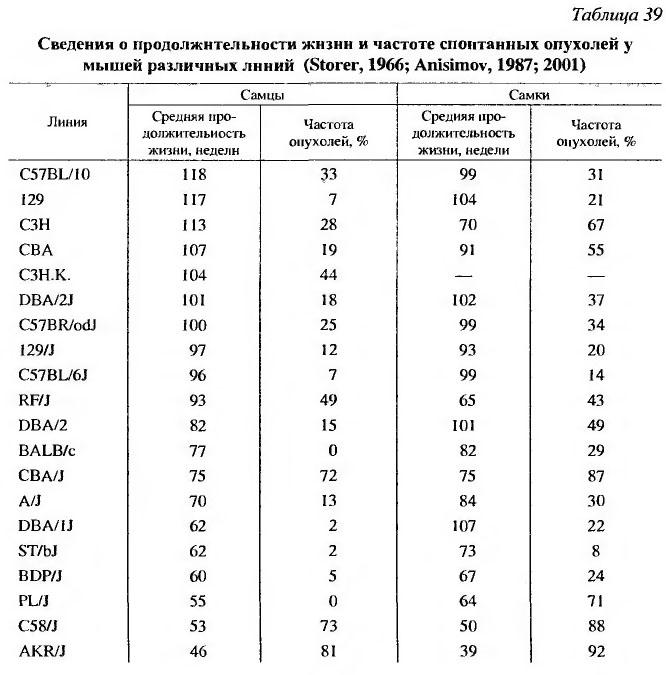
Уже отмечалось выше (см. раздел 5.2), что не наблюдается положительной корреляции между продолжительностью жизни и частотой развития опухолей, выявляемых у инбредных мышей различных линий (табл. 39) или животных одной линии, но разных популяций (Anisimov, 1987, 2001).
Отсутствует и корреляция между видовой продолжительностью жизни и частотой рака. Так, расчеты показывают, что у человека с продолжительностью жизни 70 лет, крысы (2.5-3 года) и мыши (2 года) кумулятивная частота новообразований составляет 30 % (Anisimov, 1987). Однако при нормализации по количеству клеток в организме оказывается, что мышь более склонна к развитию опухолей, чем человек (Miller, 1991). Этот феномен склонны объяснять тем обстоятельством, что в соматических клетках мыши теломераза более экспрессирована, чем в клетках человека, чему соответствует значительно большая длина теломер у мыши по сравнению с человеком (Wright, Shay, 2000). Вместе с тем видовая продолжительность жизни млекопитающих хорошо коррелирует с эффективностью систем репарации ДНК (Cutler, 1991) и резистентностью их клеток к окислительному стрессу, вызываемому различными агентами (Каpahi et al., 1999). Эффективность репарации алкилированного канцерогенными нитрососоединениями в О6-положении гуанина в ДНК человека в сотни раз выше, чем у мыши, что соответствует большей резистентности человека к этим агентам (Likhachev, 1990). Показано, что имеет место высокая положительная корреляция между эффективностью репарации вызываемых канцерогеном бензо(а)пиреном повреждений в ДНК различных органов и продолжительностью жизни долгоживущих мышей линии C57BL/6 и короткоживущих мышей BALB/c (Boerrigter, Wei, 1995). Анализ данных по частоте рака у генетически модифицированных животных с увеличенной продолжительностью жизни (см. главу 6) свидетельствует о снижении у них частоты злокачественных новообразований.
Установлено, что ограничение калорийности питания практически на всех биологических объектах сопровождается увеличением продолжительности жизни (Wendruch, Walford, 1988; Roth et al., 1999). Ключевым биологическим параметром при этом является низкий уровень инсулина и IGF-1. У нематод и плодовых мушек были выявлены мутационные изменения генов в системе передачи сигнала от инсулинового рецептора к транскрипционному фактору daf-16, которые ассоциированы с существенным увеличением продолжительности жизни. Все описанные мутации (age-1, daf-2, СHIСО, InR и др.) находятся в генах, предшествующих daf-16 в инсулиновом каскаде, daf-16 является транскрипционным фактором и оказывает свое действие, связываясь в промоторных областях генов, регулируемых инсулином (insulin-response elements, IRE). Изучение возрастного распределения частот аллелей Апо CIII, один из которых располагается в 5'-нетранслируемой области этого гена (Т-455С) в пределах функционального участка связывания инсулина, показало, что частота мутационного события в IRE гена Апо СIII человека строго положительно коррелирует с продолжительностью жизни (Anisimov S. et al, 2001). Таким образом, впервые представлены доказательства, что мутация в системе передачи сигнала инсулина, находящаяся ниже гена daf-16, непосредственно связана с долголетием (Schwartz et al., 2002). Гиперинсулинемия может способствовать окислительному стрессу и тем самым независимо от гипергликемии ускорять старение и формирование ассоциированных с возрастом заболеваний, таких как сахарный диабет, атеросклероз, гипертоническая болезнь и рак, Гиперинсулинемия развивается вторично в связи с нарушенной способностью инсулина стимулировать метаболизм глюкозы в скелетных мышцах (резистентность к инсулину). Другой способствующий старению эффект инсулина состоит в стимуляции полиненасыщенных жирных кислот и угнетении протеосома (Facchini et al., 2000). Авторы полагают, что данные о существенном увеличении продолжительности жизни С. elegans с мутациями, в частности в гене daf-2, тормозящими передачу сигнала инсулина (Kimura et al., 1997), или увеличении продолжительности жизни при ограничении калорийности питания, снижающем уровень глюкозы и инсулина в крови (Lane et al., 1995) и окислительный стресс (Xu, Badr, 1999), могут служить подтверждением их гипотезы. Аналогичным образом Matsutnoto и соавт. (2000) связывают гипоталамические нарушения и гиперинсулинемию с ускоренным старением и нарушением регуляции репродуктивной функции, энергии и веса тела. Снижение уровня гормона роста, инсулина и IGF-1 - ведущие факторы увеличения продолжительности жизни у карликовых мышей Эймса (Bartke et al., 2001). Доказано, что у столетних существенно реже наблюдается резистентность к инсулину и сохранена функция -клеток инсулярного аппарата, чем в более молодых возрастных группах (Paolisso et al., 2001) и снижена частота рака (Pompei et al., 2001; Bordin et al., 1999). В ряде недавних работ резистентность к инсулину и гиперинсулинемия рассматриваются как новые важные факторы в развитии рака (Colangelo et al., 2002; Gupta et al., 2002), причем указывается, что разработка лекарственных средств, восстанавливающих чувствительность к инсулину и соответственно снижающих уровень инсулина, может стать наиболее приоритетным направлением в профилактике рака (Gupta et al., 2002).
Замедление старения ограничением калорийности питания у мышей, крыс и обезьян сопровождается торможением спонтанного канцерогенеза (Weindruch, Walford, 1988; Roth et al., 1999), тогда как избыточный вес является фактором риска рака и ряда других ассоциированных с возрастом заболеваний (Berstein, 1997). В основе этого эффекта, как полагают, лежит снижение оксидативного стресса, снижение концентрации глюкозы в крови и уменьшение неэнзиматического присоединения глюкозы к долгоживущим белкам, например к гемоглобину (Masoro, 2000; Ulrich, Cerami, 2001). Снижение концентрации глюкозы приводит к снижению как гликозилирования белков, так и перекисного окисления липидов. Определяющим негативный эффект гликозилирования является не собственно присоединение глюкозы к долгоживушим белкам, а происходящее вследствие этого обусловленное свободными радикалами их окислительное повреждение. Нуклеотиды и ДНК также подвергаются неэнзиматическому гликозилироваиию, что приводит к мутациям из-за прямого повреждения ДНК и инактивации систем репарации ошибок рекомбинации, это также вызывает повышенную ломкость хромосом. В настоящее время изучаются подходы к предупреждению влияния гликозилирования на долгоживущие белки с помощью фармакологических и генетических воздействий. Использование миметиков калорийно ограниченной диеты, повышающих чувствительность к инсулину и снижающих уровень глюкозы в организме, рассматривается как перспективное направление в современной геронтологии (Mattson et al., 2002). Длительное введение мышам и крысам антидиабетических бигуанидов приводило к замедлению старения репродуктивной системы, увеличению продолжительности жизни животных и снижению у них частоты развития новообразований (Oilman, Anisimov, 1980; Anisimov, 1987). Применение различных геропротекторов, то есть средств, увеличивающих продолжительность жизни, по-разному влияло на развитие новообразований, что определялось в основном типом замедления старения в популяции, подвергшейся такому воздействию (Anisimov, 2001) (см. главу 10).
Наши наблюдения свидетельствуют о том, что более "прямоугольный" характер кривых выживания ассоциирован с увеличенной скоростью развития фатальных опухолей у крыс. С другой стороны, увеличение в популяции животных доли слабых, уязвимых (frailty) особей в молодом возрасте, приводит к уменьшению показателей смертности в старости и соответственно уменьшению скорости развития фатальных опухолей (Anisimov, 1987, 1998). Принимая в расчет экспоненциальный характер отношения между частотой возникновения опухолей и возрастом, мы рассчитали коэффициент корреляции между параметрами популяционного старения и развитием опухолей в десяти группах интактных крыс, находившихся в одинаковых условиях и использованных в качестве контрольных животных в наших экспериментах (Anisimov, 1987). Для каждой группы строились таблицы дожития, рассчитывались показатели смертности (R), средней продолжительности жизни и вычислялась константа а в соответствии с уравнением Гомпертца ( , где Ro - смертность к моменту времени (t) = 0; = константа). Для каждой группы рассчитывалась также кумулятивная частота опухолей к 1000-му дню жизни животных в соответствии "life-table" методом и по уравнению: , где Q = общее число опухолей, t = время, Q = Qo к моменту времени t = 0 и к = константа, - определялась величина этой константы. Была установлена высокая положительная корреляция между скоростью старения популяции (оцененная как ) и кумулятивной частотой развития опухолей Q (r = 0.70, р < 0.05) и константой к (r = 0.77, р < 0.05). Очевидно, что константа k может рассматриваться как показатель скорости увеличения с возрастом частоты опухолей в популяции. Таким образом, наши данные свидетельствуют о положительной корреляции между появлением опухолей, скоростью возрастного увеличения частоты опухолей и скоростью старения самой популяции (Anisimov, 1987, 1998). К аналогичному выводу привел анализ результатов воздействия различных геропротекторов на кривые смертности мышей и крыс и кривые возникновения в этих же популяциях частоты новообразований (Anisimov, 1987, 1998, 2000).
Следует отметить, что с периода начала индустриальной революции в конце XVIII века и вплоть до середины XX века в экономически развитых странах наблюдалась прогрессивная "ректангуляция" кривой Гомперца, которой соответствовало постепенное увеличение заболеваемости раком (Totter, 1980; Parkin et al., 2001). Частота всех злокачественных новообразований в США и многих других странах, включая Россию, существенно увеличивалась с 1960 по 1990 г. как у мужчин, так и у женщин, что коррелировало с увеличением продолжительности жизни. Начиная с 90-х годов прошлого века, в наиболее экономически развитых странах, таких как США, Швеция и Дания, отмечается замедление и даже снижение нарастания частоты рака (Ries et al, 2001). Следует отметить, что именно во второй половине XX века в наиболее развитых странах изменился характер траекторий смертности - ректангуляция сменилась их параллельным сдвигом, что сопровождалось уменьшением смертности в самых старших возрастах (Vaupel et al., 1998; Yashin et al., 2002). В то же время в менее экономически развитых странах, например в России, частота злокачественных новообразований продолжает увеличиваться. Очевидно, что увеличение продолжительности жизни и характера траектории смертности по типу параллельного сдвига или даже увеличения ее наклона, будет способствовать снижению частоты злокачественных новообразований.
См. также:
 5.1. Введение. Гипотезы о взаимосвязи рака и старения
5.1. Введение. Гипотезы о взаимосвязи рака и старения
5.2. Возрастное увеличение частоты спонтанных опухолей
5.3. Чувствительность к канцерогенам в разном возрасте
5.4. Старение и многостадийная модель канцерогенеза
5.5. Влияние старения на рост перевиваемых опухолей
5.6. Клеточное старение и канцерогенез: роль теломер и теломеразы
5.7. Канцерогенное старение
5.8. Ускоряет ли преждевременное старение развитие опухолей?
5.9. Окислительный стресс, старение и канцерогенез
5.11. Заключение
 Обсудить на форуме
Обсудить на форуме

