Утром я была я или уже не я?
Никакой разницы не припомню.
Но если уж я не я, то кем меня подменили?
Льюис Кэрролл. Алиса в Стране чудес
Известно, что с возрастом учащаются случаи различных инфекционных заболеваний, аутоиммунных процессов и опухолей. Возможно, это частично обусловлено возрастными дефектами иммунной системы. Связь столь широкого круга зависимых от возраста патологических процессов с дефектами иммунной системы привела к появлению предположения, что старение иммунной системы может ограничивать продолжительность жизни (Walford, 1969). Однако, несмотря на то что выполнено множество экспериментальных и клинических исследований, свидетельствующих о возрастном истощении иммунной системы, имеющихся данных все же недостаточно для объяснения всех проявлений старения (Lipschitz, 1987; Miller, 1999; Полякова и др., 2001; Романюха, Яшин, 2001; Uemura et al., 2002). Множество клеточных и гуморальных компонентов, вовлекаемых в иммунные реакции (табл. 24) и большое число модулирующих неиммунных факторов, которые также могут изменяться в старости (Фрейдлин, 1998; Hadden, 1998) не позволяют и сегодня нарисовать исчерпывающую картину иммуностарения.

Иммунная и гемопоэтическая системы тесно связаны, поскольку имеют единое происхождение от общих поливалентных стволовых клеток. Обе играют ключевую роль в защите организма, предупреждении развития опухолей и возникновении ответа на инфекционные агенты (Lipschitz, 1987). Однако оказывается, что с возрастом основной гемопоэз у животных и у человека не изменяется или изменяется минимально (Lipschitz, 1987). Резервные возможности могут сужаться, что приводит к снижению способности реагировать на стрессорное воздействие. Периферические лимфоидные органы, такие как селезенка и лимфоузлы, с возрастом не претерпевают закономерных изменений в размерах. Возраст не вызывает каких-либо поражений костного мозга. Продукция стволовых клеток, как правило, хорошо сохранена в старом возрасте, хотя и имеются данные о слабых изменениях скорости их репликации (Harrison et al., 1978).
Считается, что инволюция тимуса, начинающаяся при половом созревании, является главным возрастным изменением иммунной системы. Такая инволюция состоит в прогрессивной потере клеточности с истощением лимфоидного пула клеток в зонах коры и кистозными изменениями эпителиальных клеток. Они являются источником различных пептидов, вовлекаемых в дифференцирующиеся лимфоидные клетки (Т-клетки) из более молодых лимфоидных клеток. Выход дифференцированных Т-клеток снижается с увеличением возраста (Globerson et al., 1990). Прогрессивно снижаются синтез и секреция полипептидных гормонов тимуса, таких как тимозин, тимопоэтин и тимулин. Считается установленным, что снижение эндокринной активности тимуса играет ключевую роль в возрастных дисфункциях иммунной системы, поскольку заместительная терапия введением гормонов способна восстановить различные иммунные функции в старости (Zatz, Goldstein, 1985). Обмен цинка, который играет существенную роль в иммунокомпетенции, в старости снижается, тогда как добавки цинка могут восстановить иммунные функции (Fabris et al., 1990).
Зрелые Т-клетки, лимфоциты (В-клетки) костного мозга и естественные клетки-киллеры (NK-клетки) могут быть определены в крови и лимфоидных органах с помощью специфических моноклональных антител. У человека с помощью этого метода не выявлено серьезных изменений соотношения различных субпопуляций лимфоцитов. Однако обнаружены серьезные изменения функционирования Т-лимфоцитов (Thompson et al., 1987). В то время как общее количество Т-клеток в периферической крови в старости заметно не изменяется, наблюдаются четкие различия в относительном количестве подтипов Т-клеток (Thompson et al.,1987; Lipschitz, 1987; Uemura et al., 2002). Количество незрелых лимфоцитов Т-предшествен никои увеличивается с возрастом, так же как и процент частично активированных Т-лимфоцитов. несущих маркеры незрелого фенотипа тимуса. Имеет место относительное увеличение цитотоксических супрессорных Т-клеток и уменьшение количества хелперов/индукторов Т-клеток. Функциональные дефекты клеточно опосредованного иммунитета коррелируют с уменьшением популяции хелперов/индукторов (Upschitz, 1987; Thompson et al., 1987, Uemura et al., 2002). Клетки, полученные от старых людей или лабораторных животных, менее способны к ответу на аллогенные лимфоциты, фитогемагглютинин, конканавалин А и растворимый антиген. Лимфоциты более старых мышей обладают меньшей способностью вызывать реакции отторжения, чем те, которые получены от более молодых особей тех же инбредных линий (Thompson et al., 1987).
Половина здоровых людей в возрасте старше 50 лет страдает кожной гиперчувствительностью (Lipschitz, 1987; Дильман, 1987). Уменьшение количества хелперов/индукторов Т-клеток и функций клеточно опосредованного иммунитета сопровождается ростом количества антител и аутоиммунных реакций (Thompson et al., 1987).
Труднее выявить возрастные изменения гуморального иммунитета (функция В-клеток). Исследования влияния возраста на продукцию антител дают противоречивые результаты, возможно, из-за широкой вариабельности этих показателей, характерной для стареющих индивидуумов. Однако твердо установлено, что старение значимо ассоциируется с присутствием различных антител, особенно антител против ядерных антигенов. Получены также доказательства, что старение влияет на скорость продукции антител активированными В-клетками (Lipschitz, 1987; Uemura et al., 2002).
Что касается функциональных изменений, то их нарушения отмечены на различных уровнях. Во-первых, способность к пролиферации Т-клеток старых индивидуумов, как правило, снижена вне зависимости от стимуляции (антигенами, митогенами), и дефект касается как снижения числа клеток, отвечающих на стимуляцию, так и преждевременного истощения мощности клона отвечающих клеток.
Во-вторых, подавляется ответ на многие интерлейкины, которые физиологически опосредуют модуляцию пролиферативной реакции. Этот феномен был зарегистрирован не только в отношении Т-клеток, но также и для NK-клеток, которые менее чувствительны в старом возрасте к действию интерлейкина-2 или интерферона (Provincial!, Fabris, 1990). В отношении вспомогательных клеток (фагоцитов, макрофагов) известно, что их количество и функция не изменяются с возрастом, а при определенных обстоятельствах оказывается, что их активность усиливается.
Показано, что функции тимуса с возрастом снижаются или утрачиваются в такой последовательности (Полякова и др., 2001):
- влияние на репопуляцию Т-зависимых участков лимфоузлов;
- влияние на выработку реагирующих на ФГА и конканавалин А Т-клеток селезенки;
- влияние на хелперную активность Т-клеток селезенки;
- влияние на реактивность клеток селезенки к аллогенным лимфоцитам.
Установлено, что развивающееся по мере увеличения возраста истощение иммунной системы, проявляющееся прежде всего уменьшением числа Т-лимфоцитов и нарушением дифференцировки В-лимфоцитов в продуцирующие антитела плазматические клетки, способствует развитию аутоиммунных заболеваний и злокачественных новообразований (Miller, 1999; Полякова и др., 2001; Романюха, Яшин, 2001). Снижению иммунной функции Т-клеток способствуют такие факторы, как дефекты стволовых клеток, инволюция тимуса, дефекты в вырабатывающих антигены клетках, старение покоящихся иммунных клеток, репликативное истощение клонально размножающихся клеток (Pawelec et al., 1999; Полякова и др., 2001). Подчеркивается, что одним из наиболее значимых возрастных нарушений в популяции Т-клеток является изменение соотношения долей естественных Т-клеток, которое уменьшается, и долей, обладающих иммунной памятью сопутствующих Т-клеток, которое увеличивается. Возрастная инволюция тимуса сопровождается снижением секреции гормонов тимуса и продукции множества медиаторов иммунного ответа, таких как тимопоэтины, тимозин -1, тимулин, тимический гуморальный фактор и др. (Полякова и др., 2001) (рис. 12).
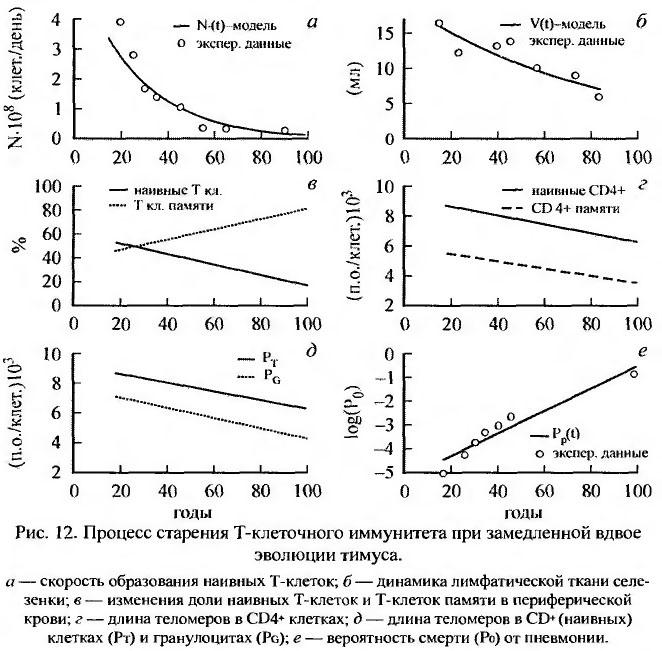
Следует отметить, что у лиц, проживших более 100 лет и сохранивших относительное здоровье, уровень ряда иммунологических показателей соответствует показателям 50-60-летних (Franceschi et al., 1995). Результаты математического моделирования возрастной динамики иммунологического старения показали, что среднее время жизни Т-клеток памяти с возрастом увеличивается, а их способность реагировать на антигены снижается (Романюха, Яшин, 2001). Уменьшение антигенной нагрузки организма повышает его резистентность в среднем возрасте. Замедление инволюции тимуса, с одной стороны, обеспечивает резистентность к новым инфекциям в пожилом возрасте, но, с другой - приводит к значительному уменьшению силы и продолжительности иммунной памяти. Снижение репликативного потенциала клеток-предшественников Т-лимфоцитов рассматривается как фактор, обусловливающий снижение эффективности методов иммунокоррек-ции в старших возрастах (Романюха, Яшин, 2001).
Важным достижением в изучении механизмов иммуностарения стало установление роли в его развитии возрастных изменений в нервной и эндокринной системах. Связь между нервной и иммунной системами опосредуется гормонами и нейромедиаторами, которые достигают лимфоидных органов и клеток через кровь или прямые связи с вегетативной нервной системой (Hadden, I998). Нейроэндокринно-иммунные взаимодействия осуществляются циркуляцией гуморальных факторов эпифизарно-гипоталамо-гипофизарной системы, либо непосредственно нейропептидами и гормонами, либо опосредованно через действие компонентов этой системы на секрецию гормонов периферических эндокринных желез, которые также обладают иммуномодулирующей активностью (Hadden, 1998). В экспериментах было продемонстрировано, что воздействие на старых животных гормонами щитовидной железы, гормоном роста и аналогами гормона, высвобождающего ЛГ, способно индуцировать реактивацию эндокринной функции тимуса, восстанавливать различные связанные с возрастом периферические иммунодефициты, такие как функциональная полноценность Т-клеток, циклоксичность NK-клеток (Fabris, 1991).
По-видимому, нейроэндокринная система действует не только как модулятор иммунной системы, но также как мишень для сигналов, генерируемых в иммунной системе. Примерами подобных взаимодействий являются изменения нейроэндокринного баланса, которые могут быть индуцированы удалением соответствующих лимфоидных органов (например, тимуса) или нарушением функции иммунной системы в результате реагирования на иммуногенные или толерогенные дозы антигена. Кроме того, зрелые лимфоидные клетки, стимулируемые антигеном, продуцируют гуморальные факторы, сходные (если не идентичные) с классическими гормонами и нейромедиаторами (такими, как АКТГ, ТТГ, гормон роста, пролактин, гамма-адреналины). Эти реципрокные связи между нейроэндокринной и иммунной системами имеют место в течение всей жизни, но приобретают особое значение в период старения (Fabris, 1991).
В последние годы было установлено, что некоторые иммуномодуляторы, в частности витамин Е, калорийно ограниченная диета и физические упражнения, вакцины, пептидные препараты тимуса могут восстанавливать компетентность иммунных клеток в старом организме и увеличивать продолжительность жизни животных (Morozov, Khavinson, 1997; Anisimov et al., 2000, 2001a; Коркушко и др., 2002; Hirokawa, Utsuyama, 2002) (см. главу 9).
См. также:
 4.1. Возрастные изменения нервной системы и старение организма
4.1. Возрастные изменения нервной системы и старение организма
4.2. Стресс и старение
4.4. Система гормон роста-инсулинподобный фактор роста-1 (1GF-1): роль в старении и долголетии
4.5. Нейроэндокринологическая (элевационная) теория старения и формирования возрастной патологии
4.6. Эпифиз и механизмы старения
 Обсудить на форуме
Обсудить на форуме

