Использовав регрессионный анализ данных о скорости укорочения теломер в клетках человека из 15 различных тканей и органов, Takubo et al. (2002) установили, что они в среднем укорачиваются на 20-60 пар оснований в год. Авторы подчеркивают, что длина теломер не имеет отчетливой корреляции со временем обновления клеток in vivo и скорее является индивидуальной характеристикой.
Было установлено, что при введении теломеразы в клетки фибробластов человека, которые в норме делятся лишь 50-80 раз, они способны поделиться 280 раз без каких-либо признаков старения и патологии. Тщательное исследование показало отсутствие в этих клетках таких признаков малигнизации, как нестабильность хромосом, независимый от добавления натуральной сыворотки рост, отсутствие контактного торможения и потеря контроля клеточного цикла (Bodnar et al., 1998; Morales et al., 1999) и, что особенно важно, из них не развиваются опухоли при трансплантации бестимусным мышам (Blasco et al., 1997; Jiang et al., 1999). Полученные данные (табл. 18) свидетельствуют о том, что экспрессия теломеразы в культуре клеток человека совсем не обязательно вызывает развитие рака, то есть лишена свойств онкогена, которые ей приписывали.
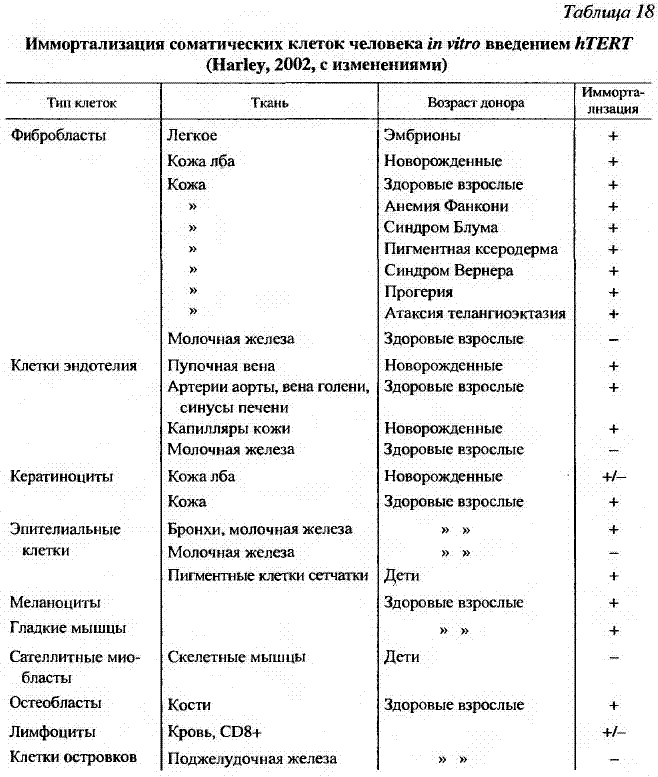
Видимо, основным свойством теломеразы является контроль клеточного деления, а для возникновения опухолевого роста необходимы дополнительные мутации и факторы.
Доказательства правомочности такого предположения были представлены Т. Kiyono и соавт. (1998), которые показали, что введения каталитического компонента теломеразы hTERT или теломеразной активности с помощью онкобелка вируса папилломы человека Е7 в кератиноциты или клетки эпителия человека недостаточно для их полной иммортализации. Она наступает лишь при дополнительном торможении регуляции антионкогена Rb/pl6 или при угнетении экспрессии р16 в качестве второй важнейшей ступени этого процесса. Элиминация антионкогена р53 таким свойством не обладала. С другой стороны, протоонкоген с-Мус может активировать экспрессию теломеразы (Wang et al., 2000). С помощью опосредованного микроклетками переноса маркированную геном nео хромосому 20 из стареющих и молодых диплоидных фибробластов человека ввели в молодые фибробласты. Во всех новообразованных клонах наблюдалось уменьшение пролиферативного потенциала на 17-18 удвоений популяции (Егоров, 1997). Авторы склонны рассматривать полученные данные как свидетельство того, что отдельные теломеры способны ограничить пролиферативный потенциал клеток.
В. М. Михельсон (Mikhelson, 2001), критически рассмотрев имеющиеся данные о роли укорочения теломер в старении, полагает, что наблюдаемые противоречия обусловлены репликативным мозаицизмом клеток. Он полагает, что истощение пролиферативного потенциала клеток в некоторых участках тканей может быть достаточно для возникновения ассоциированных со старением заболеваний. Комбинация различных нарушений, постепенно нарастающих с возрастом, и представляет собой старение, что противоречит точке зрения Л. Хейфлика, который считает необходимым различать связанные с возрастом заболевания и "чистое" старение.
Т. von Zglinicki (2002) рассматривает гетерогенность репликативного старения как один из существенных моментов этого феномена, причем значительная часть клеток перестает делиться уже после нескольких удвоений, тогда как другая продолжает делиться даже после прохождения "предела" Хейфлика для популяции в целом. Математическое моделирование этого процесса позволило предположить, что быстро стареют и прекращают делиться клетки с наиболее короткими теломерами, и скорость укорочения теломер может быть выше в рано стареющих клетках клона (Arino et al., 1995). Следует отметить, что этот вывод еще не подтвержден экспериментально. Более того, преждевременное старение может быть индуцировано воздействием самых разнообразных стрессоров, не вовлекающих в этот процесс укорочение теломер (von Zglinicki, 2002).
В работах, специально посвященных вопросу о роли стресса в ре-пликативном старении клеток (Saretzki, von Zglinicki, 2002; von Zglinicki, 2002) приведены убедительные доказательства укорочения теломер, сопровождающегося укорочением продолжительности жизни клеток in vitro, при воздействии мягких стрессоров, например хронической гипоксии, воздействии гомоцистеина, низких доз tert-бутилгидропероксида или перекиси водорода. Была обнаружена существенная обратная корреляция между скоростью укорочения теломер и антиоксидазной активностью при исследовании этих показателей в различных линиях фибробластов человека (Saretzki, vonZglinickJ, 2002). Фибробласты с низкой активностью антиоксидазных систем укорачивали свои теломеры быстрее, чем фибробласты с высокой антиокислительной активностью. В фибрибластах человека с низкой активностью этих систем, трансфецированных геном супероксиддисмутазы, замедлялось укорочение теломер и увеличивалась продолжительность жизни этих клеток в условиях как в норме, так и при гипероксии. Подчеркивается, что антиоксиданты не только останавливают ускоренное укорочение теломер, вызванное окислительным стрессом, но и увеличивают продолжительность репликативной жизни клеток, замедляя процесс укорочения теломер (von Zglinicki, 2002). Предполагается, что укорочение теломер in vivo при некоторых заболеваниях, например сосудистой деменции, атеросклерозе или апластической анемии, может быть обусловлено оксидатив-ным стрессом, играющим важную роль в их патогенезе. В случае, если это предположение подтвердится, то длина теломер может оказаться хорошим биомаркером кумулятивного воздействия стресса и прогностическим показателем риска заболеваний в пожилом возрасте (von Zglinicki, 2002). Таким образом, может быть, что укорочение теломер является не только счетчиком клеточных делений, но и мутационного процесса, поскольку короткие теломеры запускают старение в ответ на окислительный стресс и нерепарированные мутации. По образному выражению Т. von Zglinicki (2002), теломеры действуют как "сторожа" клеток и при повреждении генома удаляют "опасные" клетки из пула пролиферирующих клеток (рис. 10).

Вместе с тем показано, что старение некоторых тканей, например эпителиальных клеток слизистой полости рта или роговицы глаза человека in vivo, не сопровождается укорочением теломер (Egan et al., 1998). Экспрессия белка аденовируса 12 Е1В 54К в нормальных клетках человека сопровождалась существенным увеличением их пролиферативного потенциала (до 100 удвоений). Когда затем деления все же прекратились и клетки перешли в фазу старения, то какого-либо существенного укорочения их теломер выявлено не было (Gallimore et al., 1997). Изучение длины теломер в ДНК различных тканей у людей в возрасте от новорожденных до столетних показало, что она уменьшается на 29-60 пар оснований (п. о.) в год в печени, коре почек и селезенке, но не в коре мозжечка или миокарде (Takubo et al., 2002). Наиболее длинные теломеры наблюдались в миокарде и наиболее короткие - в печени или корковом слое почек. Авторы не обнаружили отчетливой корреляции между длиной теломер и временем обновления тканей in vivo и полагают, что длина теломер является скорее индивидуальной характеристикой. Так, в быстро обновляющихся тканях, например в слизистом эпителии желудочно-кишечного тракта, величина укорочения теломер на одно клеточное деление меньше, чем, например, в медленно обновляющемся эпителии печени или почек (табл. 19).
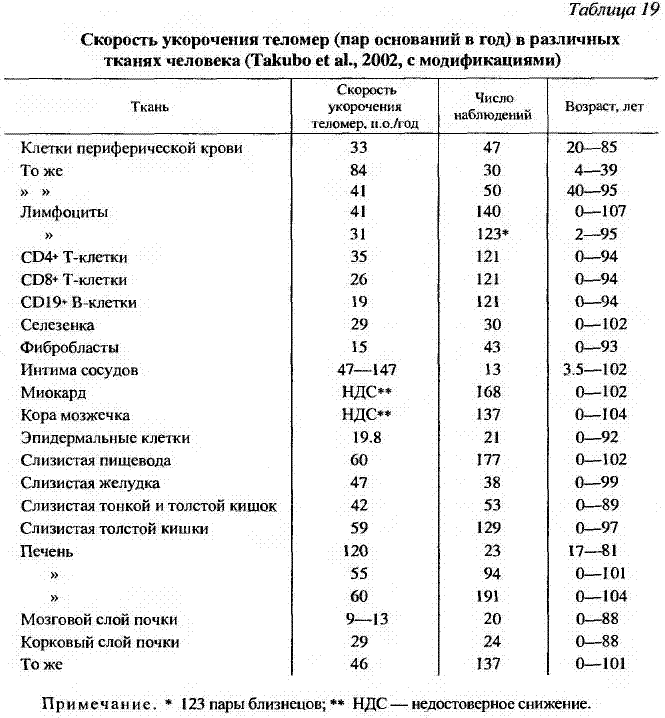
Важным наблюдением оказалась высокая степень корреляции между длиной теломер в разных органах отдельных индивидуумов. Так, у лица с большой длиной теломеры в одном органе были длинные теломеры и в других органах. У столетних средняя длина теломер (примерно >8.7 т. п. о. в печени) для пяти органов была всегда большей, чем в фибробластах или эпителиальных клетках, культивируемых in vitro (Takubo et al., 2002). Длина теломер в мононуклеарах крови столетних была короче, чем в более молодых возрастных группах, и у женщин была больше чем у мужчин (Tauchi et al., 1999).
Экспрессию активности теломеразы наблюдали в печени крыс после частичной гепатэктомии (Tsujiuchi et al., 1998), то есть в процессе регенерации. Не удалось наблюдать существенных изменений в продолжительности жизни или развитии мышей с "выключенным" (knockout) геном теломеразы (Lee et al., 1998).
Важно подчеркнуть, что in vivo стволовые клетки не имеют репликативного лимита. Расчеты, основанные на данных о длине клеточного цикла, показали, что герминогенные клетки эпителия языка мыши подвергаются 565 удвоениям в течение 2 лет жизни (Cameron, 1972), а стволовые клетки тонкой кишки мыши проходят от 820 до 2200 удвоений в течение жизни без каких-либо признаков клеточной гибели (Potten, Loeffler, 1990) (табл. 20).

В блестящем обзоре, посвященном проблеме клеточного старения in vitro и in vivo, Rubin (1997) пришел к выводу, что нет оснований полагать, что лимит Хейфлика имеет место в самообновляющихся тканях in vivo и играет какую-либо роль в старении организма. Реально наблюдаемой характеристикой старения in vivo является уменьшение интенсивности клеточной пролиферации в большинстве тканей животных (Anisimov, 1987; Baserga, 1977; Rubin, 1997, 2002). Отмечается увеличение длительности клеточного цикла и его вариабельности с возрастом.
Вопрос о соответствии клеточного старения и старения тканей in vivo был специально рассмотрен P. J. Homsby (2002). Он разделяет точку зрения, согласно которой клеточное старение является феноменом, имеющим место только в культуре клеток и не может рассматриваться как адекватная модель старения in vivo. Основные выводы, к которым пришел автор, следующие.
Во-первых, укорочение длины теломер, которым опосредуется клеточное (репликативное) старение, наблюдается во многих, тканях человека при старении. Длина теломер в большинстве тканей человека коротка потому, что в них отсутствует теломераза, необходимая для поддержания их длины. Отсутствие теломеразной активности определяется отсутствием экспрессии каталитической субъединицы теломеразы (TERT) рибонуклеопротеидного комплекса теломеразы.
Во-вторых, неясно, приводит ли именно укорочение теломер к клеточному старению или некоторым другим ситуациям, в которых может оказаться клетка, например кризису.
B-третьих, накапливается все больше данных, свидетельствующих о том, что клетки грызунов не являются адекватной моделью клеточного старения человека. Выявлены существенные различия в биологии теломер мыши и долгоживущих видов (например, человека или коровы).
В-четвертых, необходимо более глубокое изучение старения тканей человека для выявления старых клеток в этих тканях, разработка новых экспериментальных подходов к исследованию последствий укорочения теломер.
При делении клетки в отсутствие теломеразы недореплицируется примерно 40-100 т. п. о. После того как нормальная клетка человека поделилась определенное число раз, которое зависит от типа клеток и условий культивирования, ее теломеры укорачиваются настолько, что они запускают остановку клеточного цикла, приводя клетку в терминальное неделящееся состояние. Именно его называют "клеточным старением". Однако в настоящее время появились доказательства того, что клетка может переходить в биохимически подобное состояние, не обязательно связанное с укорочением теломер. Поэтому в настоящее время под клеточным старением понимают процесс, который побуждает клетку к переходу в это "старое" состояние вне зависимости от того, вовлечено в этот процесс укорочение теломер или нет (Hornsby, 2002). Другой часто используемый термин - "репликативное старение" - более определенно связан с клеточным старением, обусловленным укорочением теломер (обозначается также как M1). При репликативном старении дальнейшее деление клетки блокируется такими ингибиторами клеточной пролиферации, как p21SDH/WAF/CIP1 и p161NK4A (Smith, Pereira-Smith, 1996; Marcotte, Wang, 2002). В тех случаях, когда репликативное старение прерывается онкобелком, например антигеном SV4O Т, клетки ускользают из этого состояния и переходят в другое, называемое кризисом, или М2. В этом состоянии самые короткие тело меры подвергаются слиянию конец-в-конец и циклам разрывов и слияний хромосом, которые переводят клетки в состояние апоптоза, что приводит к гибели значительной части клеток, не наблюдающейся при репликативном старении (Ml).
Клетки в культуре могут переходить в состояние клеточного старения под влиянием различных факторов окружающей среды, таких как окислительный стресс, ионизирующая радиация или эктопическая секреция некоторых сигнальных молекул и циклин-зависимых ингибиторов киназы (Marcotte, Wang, 2002). Поскольку эта воздействия могут вовлечь клетку в процесс старения без укорочения теломер, данный процесс следует отличать от его следствия - репликативного старения. Поэтому вопрос о клеточном старении в целом может быть разделен на несколько отдельных вопросов: наблюдается ли укорочение теломер in vivo и приводит ли оно к репликативному старению; если в тканях действительно формируются старые клетки, то как долго они могут находиться в ткани и являются ли они результатом репликативного старения или какого-то иного процесса? Ответ на первые два вопроса вполне положителен, тогда как гораздо меньше доказательств существования старых, клеток в тканях in vivo (Hornsby, 2002).
Считается, что наиболее существенным доказательством накопления по мере старения in vivo старых клеток в тканях является обнаружение окрашивающихся SA- gal+ (галактозидазой) клеток в коже человека, и у макак-резус - в пигментом эпителии сетчатки и в эпидермисе. В этих тканях имеет место пропорциональное возрасту донора увеличение числа клеток (Hornsby, 2002). Однако механизм возникновения таких клеток остается неясным. Неизвестно также, в какой мере изменения в экспрессии генов в этих клетках in vivo соответствуют изменениям в экспрессии, наблюдаемым при репликативном старении in vitro. Полагают, что убедительные доказательства, что старые клетки накапливаются в тканях организма с возрастом, до настоящего времени отсутствуют (Тrоеn, 2003). Выяснение этих вопросов представляется весьма важным, поскольку предполагается, что эти изменения могут приводить к формированию микроокружения, благоприятствующего развитию новообразований (Campisi, 2001; Krtolica et al., 2001; Krtolica, Campisi, 2003). Этот аспект проблемы будет подробнее рассмотрен в разделе 5.6.
Следует подчеркнуть, что клеточное старение является универсальным процессом, развивающимся как реакция клеток млекопитающих в ответ на повреждения определенного типа, включающие укорочение теломер. С точки зрения эволюционной теории клеточное старение представляется загадочным феноменом (Hornsby, 2002). Виды повреждений, которые побуждают клетку переходить в состояние старения, весьма сходны с теми, которые запускают апоптоз. С эволюционных позиций, для организма и генома в целом вхождение клетки в апоптоз весьма целесообразно, поскольку при этом удаляется поврежденная клетка, которая могла бы передать своим потомкам потенциалы" поврежденные копии генома. Поэтому остается неясным, почему в организме, в котором ежедневно рождаются и умирают миллионы клеток, имеет место клеточное старение, а не только апоптоз. Многое в этой области еще предстоит выяснить. Тем не менее очевидно, что опыты с теломеразой открывают новые перспективы как в геронтологии, так и в онкологии для диагностики рака и, что особенно важно, для его лечения.
См. также:
 3.2.1. Теломераза и теломера: основные свойства
3.2.1. Теломераза и теломера: основные свойства
3.2.3. Теломераза и теломера; мышь против человека?
 Обсудить на форуме
Обсудить на форуме

