Один из излюбленных аргументов сторонников гипотезы генетически запрограммированного предела продолжительности жизни состоит в следующем: продолжительность жизни ограничена потому, что клетки организма могут совершить лишь ограниченное, строго определенное число делений, после чего погибают [Fries, 1980].
Следует отметить, что подобное представление имеет глубокие исторические корни, а многие противоречия в публикациях по данному вопросу можно понять, лишь обратившись к его истории. Поэтому в целях ясности изложения данная проблема будет изложена в виде краткого исторического экскурса. Кроме того, чтобы обеспечить объективность и документальность такого экскурса, мы будем вынуждены обильно цитировать первоисточники и надеемся, что читатели одобрят подобный документальный стиль изложения проблемы.
Более 100 лет назад знаменитый немецкий биолог А. Вейсманн [Weismann, 1882; 1884] постулировал, что ограниченная продолжительность жизни организмов обусловлена ограниченной способностью соматических клеток к размножению. Вейсманн полагал также, что различия в долговечности разных видов животных обусловлены тем, что соматические клетки этих видов различаются по числу воспроизводимых ими поколений клеток (клетки долгоживущих видов способны совершить больше делений). Так возникло "предположение, что продолжительность жизни связана с числом поколений соматических клеток, следующих друг за другом в течение одной жизни, и что это число так же как и продолжительность жизни отдельных клеточных поколений, дано уже в зародышевой клетке" (Вейсманн. 1914. с. 481. Вейсманн объяснял "различную продолжительность жизни животных, ставя ее в зависимость от различного числа клеточных поколений, являющегося нормой для тела различных видов" [Там же, с. 47].
Следует отметить, что на рубеже XIX-XX вв. представление Вейсманна о том, что ограниченная продолжительность жизни обусловлена истощением способности клеток к размножению было необычайно популярным и разделялось многими известными учеными того времени. Так, например, русский ученый И. Р. Тарханов писал: "...в зародышевом оплодотворенном яйце определена заранее, в силу закона наследственности, вся сумма клеточных поколений, могущих развиваться за счет созидающих сил зародышевого яйца" (Тарханов. 1891. с. 551-552). По его мнению, "число клеточных поколений, могущих развиться в течение всей жизни из зародышевого яйца ... и определяет собою долголетие, ту максимальную продолжительность жизни, до которой могут достигать разнообразные организмы" (Там же, с. 542).
Концепция Вейсманна доминировала в течение 30 лет. Поэтому сенсационными стали опровергающие эту концепцию эксперименты А. Карреля [Carrel, 1912] и А. Эбелинга [Ebeling, 1913] по культивированию фибробластов сердца цыпленка вне организма (in vitro). Эти авторы сообщили, что при соответствующих условиях клетки многоклеточного организма могут размножаться как микроорганизмы практически неограниченно (эксперимент по успешному размножению клеток продолжался целых 34 года и был прекращен лишь в 1946 г. после смерти самого А. Карреля). В результате этих экспериментов, получивших всеобщее признание и широкую известность, концепция Вейсманна была оставлена и забыта, а вместо нее утвердилось прямо противоположное представление о потенциальном "бессмертии" соматических клеток, образующих смертный организм. Поэтому причины, ограничивающие продолжительность жизни, стали искать на надклеточном физиологическом уровне, в частности на уровне гормональной регуляции.
Концепция Карреля господствовала более 40 лет, и казалось, что она победила окончательно и бесповоротно. И хотя многие исследователи неоднократно наблюдали истощение способности клеток к размножению в культуре, эти случаи было принято объяснять неудачами, которые нередко сопутствовали культивированию клеток (например, контаминация вирусами или токсичность очередной партии сыворотки крови, входящей в состав культуральной среды). Популярность концепции Карреля была настолько велика, что на многочисленные исключения из установленного "правила" (неограниченный рост культур клеток животных) просто не обращали внимания, считая их методическими артефактами [Witkowski, 1987].
Э. Свим был, по-видимому, первым исследователем, который решился на радикальный пересмотр концепции Карреля. Проанализировав результаты 336 публикаций, включая и результаты своих собственных экспериментов по длительному культивированию 23 штаммов фибробластов, взятых из нормальных тканей эмбрионов кролика и цыпленка [Haff, Swim, 1956], а также 51 штамма фибробластов человека, полученных из крайней плоти, плаценты, яичек, матки и эмбриональных тканей [Swim, Parker, 1957], Свим сделал следующие выводы принципиальной важности: "...в большинстве случаев, где наблюдается рост клеток, последние в конце концов, претерпевают неспецифическую дегенерацию" [Swim, 1959, р. 145]. "Опыт многочисленных исследователей свидетельствует, что раннее культивирование клеток обычно следует определенному сценарию, который для удобства можно разделить на три фазы. В фазе I клетки быстро пролиферируют после начальной задержки (лага) и обычно могут без труда выдержать серию пересевов. Фаза II характеризуется снижением роста до тех пор, пока этот рост обычно не прекращается вовсе и клетки в конце концов не исчезают в результате неспецифической дегенерации" [Ibid, p. 159]. "Это вначале сопровождается увеличением числа гранул в цитоплазме клеток; позднее наблюдаются дегенерирующие клетки, и их численность прогрессивно увеличивается до тех пор, пока дно флаконов не покрывается плотным слоем клеточного дебриса..." [Haff, Swim, 1956, р. 201]. Был также сделан важный вывод, что прекращение размножения клеток - это не методический артефакт, обусловленный такими факторами, как размер инокулята, токсичная среда или неспособность клеток пролиферировать на стекле [Swim, Parker, 1957]. Наконец, Свим отмечает, что "иногда распознается третья стадия - появлением активно пролиферируюших клеток в культурах, находящихся в фазе II" [Swim, 1959, р. 159]. "Следует подчеркнуть, что фазы I и II представляют собой обычную картину, в то время как фаза III - сравнительно редкое явление" [Ibid, p. 160]. Свим отметил также, что клетки в фазе III часто отличаются от исходных клеток и по морфологии, и по ростовым характеристикам.
Таким образом, неограниченная способность клеток животных к делению оказалась не правилом, а исключением, причем во многих случаях неограниченно делящиеся клетки уже мало напоминают по морфологии и ростовым характеристикам исходные нормальные клетки [Swim, 1959]. Казалось бы, этот вывод давал все основания для пересмотра концепции Карреля и к возрождению интереса к концепции Вейсманна. Однако, чтобы сделать это и разрушить в общественном сознании устоявшиеся в течение десятилетий общепринятые представления, необходимо было проявить такую исключительную активность в пропаганде своих результатов и выводов, а также такую предприимчивость, которой Свим, повидимому, не обладал. Зато нашлись другие исследователи, которые блестяще справились с этой задачей.
В 60-х и 70-х годах Л. Хейфлик опубликовал большую серию научных и научно-популярных статей (см.. например: [Hayflick, Moorhead, 1961; Hayflick, 1965; 1968; 1974; Хейфлик, 19691), посвященных результатам длительного культивирования фибробластов человека. Фактически эти статьи подтверждали результаты и выводы Свима, а также концепцию Вейсманна, однако большинство читателей, не знакомых с работами этих незаслуженно забытых авторов, восприняли публикации Хейфлика как принципиально новое научное открытие.
Примечание: Подобное восприятие статей Хейфлика было отчасти обусловлено их стилем: концепция Вейсманна о существовании предела клеточных делений излагалась без упоминания ее автора, а иэ основополагающих работ Свима лишь однажды [Hayflick, Moorhead, 1961] была вскользь упомянута одна [Swim, Parker, 1957], так что создается впечатление принципиальной новизны всех полученных результатов и их интерпретации.
Поскольку публикации Хейфлика вызвали огромный интерес и получили широкую известность, ограничимся лишь кратким изложением основных положений этих статей.
В своей знаменитой публикации [Hayflick, Moorhead, 1961, р. 600] авторы пишут: "Обычная история штамма диплоидных клеток может поэтому быть разделена на три отдельные фазы... Фаза I, или фаза раннего роста, включает период, когда клетки освободились от интактной ткани и только что расположились на стекле (первичная культура).
Примечание: Нетрудно заметить, что фаза I по классификации Хейфлика и Мурхеда соответствует начальному лагу фазы I по классификации Свима [Swim, 1959].
"Фаза I заканчивается формированием первого сомкнутого слоя, в этот момент культура готова к своему первому пересеву и вступает в фазу II. Фаза II характеризуется быстрым размножением клеток... В течение этой фазы диплоидные штаммы должны пересеваться, по крайней мере, дважды в неделю с разведением в отношении 2 или 3:1".
Примечание: Очевидно, что фаза II по классификации Хейфлика и Мурхеда соответствует фазе I после начального лага по классификации Свима.
"Фаза II длится от 2 до 10 месяцев, после чего начинает происходить клеточная дегенерация. Эта дегенерация и снижение митотической активности знаменует появление фазы III. или терминальной фазы. Она характеризуется появлением дебриса... снижением митотической активности и, следовательно, более длинным периодом времени, необходимым для образования сомкнутых монослоев".
Примечание: Фаза III по классификации Хейфлика и Мурхеда точно совпадает с фазой II по классификации Свима.
Кроме того, авторы отмечают, что "может происходить изменение... порождающее "клеточную линию", чья потенциальная продолжительность жизни бесконечна" [Hayflick, Moorhead, 1961, р. 587]. Нетрудно заметить, что это "изменение" соответствует переходу культуры в фазу III по классификации Свима. Таким образом, многие принципиальные положения этой знаменитой и широко цитируемой статьи Хейфлика и Мурхеда фактически являются повторением незаслуженно забытых выводов Свима.
На основании приведенных результатов и ряда дополнительных экспериментов был сделан вывод, что клетки человека способны совершить лишь ограниченное, строго определенное число делений, а затем гибнут. В соответствии с этой клеточной версией модели шагреневой кожи человек в течение жизни неизбежно исчерпывает заложенный в клетках потенциал делений и "момент, когда наступает эта потеря, может определять предел продолжительности жизни человека" [Hayflick, 1980, р. 42]. Действительно, "оказалось, что фибробласты эмбрионов человека в культуре делятся 50±10 раз: фибробласты, взятые от момента рождения до 20 лет, делятся 30±10 раз, а клетки, взятые у доноров старше 20 лет, делятся 20±10 раз" [Хейфлик, 1969, с. 219]. Таким образом, почти через 100 лет исследователи фактически вновь вернулись к некогда отвергнутой концепции Вейсманна [Weismann, 1882], забыв, однако, ее автора.
Получило подтверждение и положение Вейсманна о том, что клетки долгоживущих видов способны совершить больше делений. Оказалось, что эмбриональные фибробласты мыши способны удвоить свою численность in vitro всего 14-28 раз, цыпленка - 15-35, человека - 40-60, а черепахи - 72-114 раз [Hayflick, 1974].
Многочисленные публикации Хейфлика 60-х и 70-х годов были восприняты многими как научная сенсация, открывающая принципиально новый подход к выяснению механизмов детерминации продолжительности жизни. Поскольку работы Свима практически не цитировались, статьи Хейфлика получили признание как первое и окончательное опровержение мифа Карреля о неограниченной способности клеток к делению. Поскольку и концепция Вейсманна о пределе клеточных делений как основе старения также была основательно забыта, статьи Хейфлика были восприняты как принципиально новая концепция, а не как возвращение к старой, первоначально выдвинутой гипотезе. Поэтому ограниченная способность клеток к делению получила название лимита Хейфлика (см., например: [Bremermann, 1982; Walford, 1983; Juckett, 1987]), а история открытия этой закономерности нередко излагается начиная лишь с работы Хейфлика и Мурхеда [Hayflick, Moorhead, 1961].
Новая (точнее, хорошо забытая старая) концепция ограниченной способности клеток к делению в настоящее время получила такую известность и столь полное признание, что о ней можно теперь прочитать даже в "Британской энциклопедии" [The New Encyclopaedia Bntanmca, 1989, vol. 1, p 148]. "Лабораторные эксперименты показали, что клетки, взятые из сложных организмов, проходят лишь ограниченное число клеточных делений до своей гибели, подтверждая идею, что клеточные события могут вызывать старение". Работы же Хейфлика, подтверждающие эту концепцию, стали рассматриваться как пример научной революции в биологии [Witkowski, 1987]. Здесь, казалось бы, пришло самое время поставить точку в этой длинной истории со счастливым концом, однако есть серьезные основания считать, что она еще не закончилась
В 1978 г нами была проведена ревизия концепции Вейсманна-Свима-Хейфлика, в результате которой были сделаны следующие выводы [Гаврилов, Гаврилова, 1978, Гаврилов, Ягужинская, 1978].
1. Массовая гибель клеток в фазе II по Свиму или в фазе III по Хейфлику - это методический артефакт, обусловленный повреждением клеток в результате процедуры пересева (обработка клеток трипсином и их последующее суспендирование). Если же оценить вероятность гибели клеток вне периодов пересева, то оказывается, что вероятность смерти клеток не увеличивается с возрастом культуры, т.е. нет оснований говорить о старении на клеточном уровне.
2. Основное содержание феномена Свима-Хейфлика in vitro - это накопление в культуре постмитотических клеток, полноценных в метаболическом отношении. Процесс образования этих метаболически полноценных постмитотических клеток больше напоминает дифференцировку клеток, чем их повреждение и старение. К этому же выводу одновременно с нами пришли и другие исследователи [Bell et al, 1978].
3. Накопление в культуре постмитотических клеток не сопровождается уменьшением абсолютного числа делящихся клеток в расчете на всю образующуюся клеточную популяцию. Иначе говоря, происходит не исчезновение делящихся клеток, а лишь разбавление культуры постмитотическими клетками. Следовательно, нет никаких оснований говорить об ограниченной способности к делению всех клеток в культуре. Напротив, например, в культуре эмбриональных фибробластов человека имеются клетки, совершающие около 170 делений (вместо общепринятых 50) и обладающие свойствами стволовых клеток с неограниченной способностью к делению.
Теперь рассмотрим сделанные выводы несколько более подробно, для чего обратимся к анализу методики культивирования клеток, состоящей в следующем. Образец ткани (например, кожи) помещают в сосуд с культуральной средой и ждут, когда из этого первичного эксплантата часть клеток мигрирует на поверхность флакона и размножившись, образует сомкнутый монослой, покрывающий дно сосуда. Никто не знает при этом, сколько клеток мигрирует из трансплантата и сколько делений они совершают. По мнению Голдштейна и соавторов, эта процедура соответствует примерно 10 удвоениям популяции [Goldstein et al., 1975]. В случае же эмбриональной ткани легкого ее обрабатывают трипсином, и суспендированные клетки пересевают в сосуд с питательной средой [Hayflick, 1968]. После того как культура клеток образует монослой, ее пересевают, обрабатывая 0 1%-ным раствором трипсина, суспендируя и перенося желаемое число клеток в новые сосуды с питательной средой. Пересев в отношении 1 2 означает, что только половина клеток, образовавших сомкнутый монослой, пересевается в сосуд с той же поверхностью. Когда и в этом сосуде в результате деления клеток образуется монослой, считают, что произошло удвоение клеточной популяции, соответствующее в среднем одному делению клеток. Подобную операцию повторяют многократно (в случае эмбриональных фибробластов человека около 50 раз), до тех пор, пока культура не потеряет способности к быстрому восстановлению своей численности. Теперь попытаемся оценить число делений, которое совершают клетки. Известно, что через сутки после операции пересева в живых остается только 50-70% клеток "молодых" культур и лишь 25% клеток "старых" культур (эти клетки являются более крупными и сильнее всего повреждаются при суспендировании) [Гаврилов, Гаврилова, 1978, 1982].
Примечание: Такое массовое разрушение клеток при пересеве в сочетании с низкими темпами восстановления численности "старых" культур и порождало иллюзию естественной массовой дегенерации и гибели клеток описанную в ряде работ [Swim, 1959, Hayflick. Moorhead, 1961, Hayfhck, 1968]
Таким образом. 40-60 пересевов соответствуют не 40-60 удвоениям популяции, а 80-120 удвоениям, которые совершают выжившие клетки. Далее, одно удвоение клеточной популяции соответствует одному делению клеток лишь в том случае, если все клетки способны делиться. На самом же деле доля делящихся клеток сильно уменьшается при культивировании и к последнему пересеву составляет всего 10-20% [Там же]. Можно показать, что в том случае, когда только 10% клеток сохраняет способность к делению, им необходимо поделиться 10 раз, чтобы численность всей культуры удвоилась [Гаврилов, Ягужинский, 1978, Гаврилов, Гаврилова, 1982]. Если учесть это обстоятельство, то окажется, что 80-120 удвоений численности клеток соответствуют 170±30 делениям. Итак, оказывается, что 50 пересевов клеток человека соответствует не 50, а 170±30 делениям.
Самое главное, однако, состоит в том, что и это число делений оказывается не окончательным. Дело в том, что гибель клеточной культуры определяется весьма своеобразно. Оказывается, культуру клеток называют мертвой лишь потому, что в течение произвольно заданного интервала времени (обычно от одной до четырех недель) ее численность не возрастает до желаемой величины (обычно задается численность, в два-четыре раза превышающая исходную).
Примечание: В качестве примера приведем следующее определение: "Культуры считались мертвыми, когда они не могли достигнуть сомкнутого монослоя после 4 недель при еженедельной смене среды" [Goldstein, Singal, 1974, р. 360).
Более того, в "мертвых" культурах еще остается 10-20% клеток, способных делиться, поэтому культура может компенсировать естественную гибель клеток (которая, кстати, не увеличивается) и даже медленно расти [Гаврилов, Гаврилова, 1978; 1982]. Оказалось также, что абсолютное число делящихся клеток в расчете на всю образующуюся клеточную массу не уменьшается, а происходит лишь их разбавление неделящимися клетками. В результате, когда доля делящихся клеток становится близка к 10%, культура действительно не может расти столь быстро, чтобы ее считали живой. Таким образом, феномен "гибели" культуры клеток не только не эквивалентен гибели клеток, но даже не означает того, что все клетки культуры имеют ограниченную способность к делению. В результате проведенных исследований нами был сделан вывод, что в культуре клеток происходит их дифференцировка с образованием неделящихся клеток, но некоторые клетки культуры являются стволовыми и могут размножаться неограниченно [Гаврилов, Ягужинский, 1978].
Некоторые из сделанных нами выводов были одновременно и независимо подтверждены другими авторами [Belletal., 1978, р. 1160]:
"...одной из наименее документированных особенностей гипотезы старения in vitro является общепринятое утверждение о том, что клетки в фазе 3 являются нежизнеспособными и в конце концов погибают. Это утверждение не имеет никакого серьезного экспериментального подтверждения ...Наш опыт свидетельствует о том, что сам по себе пересев неизбежно приводит к потере клеток. ...Мы наблюдали выживаемость фибробластов крайней плоти (штамм 1519) в культуре в течение длительного периода после того, как клеточное деление прекращалось. Наши первые культуры клеток штамма 1519 поддерживались живыми в среде Хэма ...в течение 22 месяцев и в течение периода от года до 14 месяцев после прекращения клеточных делений ...Биохимические исследования показали, что, несмотря на то, что клетки фазы 3 отличаются в некоторых отношениях от клеток фазы 2, они, по существу, нормальны. ...Сообщения о биохимических различиях между клетками в фазе 2 и фазе 3 отражают различия в дифференцировке, а не в возрасте".
Примечание: Фазы 2 и 3 в данной статье соответствуют классификации Хейфлика, а не Сейма. Повидимому, авторы данной статьи, так же как и многие другие исследователи, просто не знакомы с основополагающими работами Сейма и поэтому его статей не цитируют.
Впоследствии и сам Л. Хейфлик совместно с Т. Матсумурой и 3. Зеррудо [Matsumura et al., 1979] подтвердил отсутствие массовой гибели клеток в фазе III по его классификации. Авторы этой работы в течение шести месяцев поддерживали "мертвые" культуры, некоторые из которых за это время в 16 раз увеличили свою численность. Никаких признаков приближающейся массовой гибели клеток обнаружить не удалось, хотя срок наблюдения за "мертвыми" культурами иногда превышал даже год (для сравнения отметим, что вся предшествующая процедура 50 пересевов занимает обычно шесть-восемь месяцев) [Hayflick, 1968].
В результате этих исследований Хейфлик изменил свое представление об ограниченной продолжительности жизни клеточных культур. Первоначально он излагал свои результаты следующим образом (рис. 54):
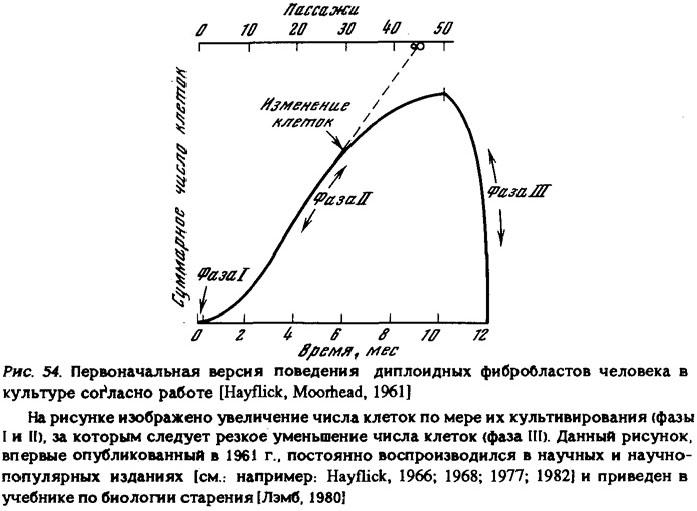
"Мы обнаружили, что фибробласты, взятые от четырехмесячного человеческого эмбриона, удваивались таким образом в числе в среднем 50 раз (от 40 до 60). Истощив свою способность к делению, клетки гибли" [Hayflick, 1968, р. 35]. В работе 1979 г. приведена уже принципиально иная схема (рис. 55)
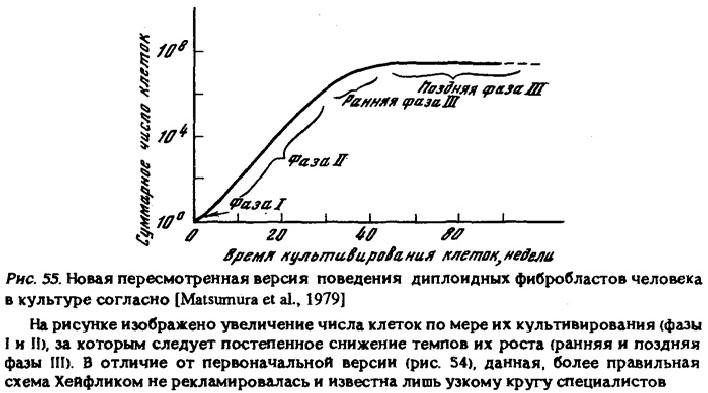
и все излагается иначе: "В ранней фазе III культура еще пролиферирует, хотя скорость пролиферации снижается. В последней фазе III клеточная пролиферация очень низка... В течение этого периода не наблюдалось ни признаков окончательной смерти культуры, ни спонтанного приобретения неограниченного пролиферативного потенциала" [Matsumura et al., 1979, р. 332, ЗЗЗ]. Следует подчеркнуть, что изменение позиции исследователя делает ему честь как ученому, поскольку цель науки состоит не в отстаивании догматов, а в поиске истины. Однако достойно сожаления то, что вскрывшаяся истина стала известна лишь очень узкому кругу специалистов, а в таких ведущих научно-популярных журналах, как "Scientific American", продолжалась реклама старой легенды, что "в конце концов клетки претерпевают различные дегенеративные изменения и гибнут" [Hayflick, 1980, р. 42]. Конечно, удивительно, что пропаганда этого мифа велась тем же автором, который годом ранее так хорошо показал его несостоятельность, однако эта загадка выходит за пределы тематики данной книги.
Примечание: Еще более удивительно, что тот же автор три года спустя опять публикует устаревшую схему (рис. 54) вместо им же исправленной схемы (рис. 55) и вновь излагает опровергнутую версию, что "после исчезновения митотической активности культура полностью дегенерирует" [Hayflick, 1982, р 641].
В то же время становится понятен механизм канонизации устаревших представлений о пределе клеточных делений в современных томах таких солидных изданий, как, например, "Британская энциклопедия".
Нетрудно заметить, что с учетом новых фактов феномен Свима-Хейфлика уже не столь очевидным образом связан с проблемой ограниченной продолжительности жизни организмов. Действительно, массовая гибель клеток могла бы приводить к гибели организма, в то время как из снижения темпов роста клеточной популяции ограниченность сроков жизни организма прямо не следует. Более того, снижение митотической активности часто является нормальным физиологическим процессом, связанным с дифференцировкой клеток. Например, при образовании нервной и мышечной ткани клетки практически теряют способность к делению, и по приведенному выше определению данные клеточные популяции могли бы быть названы мертвыми. Ясно, однако, что такое определение малоконструктивно, поскольку речь идет о высокоспециализированных и функционально полноценных клетках. Действительно, во многих случаях выполнение высокоспециализированной функции оказывается несовместимым (по крайней мере, одновременно) с клеточным делением. Известно также, что при дифференцировке клеток для переключения их метаболизма часто бывает необходимо клеточное деление, причем критический пункт в таком переключении совпадает с синтезом ДНК [Гаврилов, Гаврилова, 1978, 1982]. Таким образом, наблюдается явное сходство между счетом делений, происходящим при дифференцировке клеток, и счетом делений, наблюдаемым при их длительном культивировании. Естественным образом возникает гипотеза, что явление, обнаруженное Свимом и Хейфликом, представляет собой не какой-то принципиально новый процесс типа старения на клеточном уровне, а давно известный, хотя и малоизученный процесс дифференцировки клеток, сопровождаемый счетом делений и снижением митотической активности [Belletal., 1978; Гаврилов, Гаврилова, 1978. 1982, Гаврилов, Ягужинский, 1978]. К сожалению, относительно фибробластов человека до сих пор нельзя сказать, во что именно они дифференцируются в культуре. Возможно, что недостаточность культуральной среды по тем или иным факторам сдерживает нормальную экспрессию нового фенотипа. В принципе известно, что при определенных условиях фибробласты способны дифференцироваться в культуре с образованием неделящихся адипоцитов, однако экспрессия этого нового фенотипа может сдерживаться при недостатке ряда факторов, например инсулина, биотина и пр. Относительно же культур эпидермальных кератиноцитов человека прямо показано, что снижение темпов роста этих культур сопряжено с дифференцировкой (кератинизацией) кератиноцитов [Rheinwald, Green, 1977].
Исходя из гипотезы дифференцировки, можно ожидать, что число удвоений клеточной популяции вовсе не является фундаментальным внутренним свойством клеток, как полагает Хейфлик, а может быть сравнительно легко увеличено с помощью ингибиторов дифференцировки (некоторых факторов роста, опухолевых промоторов
Примечание: В данном конкретном случае речь идет не о трансформации клеток с помощью опухолевых промоторов а об их использовании в качестве ингибиторов дифференцировки клеток.
и т. д. ) Действительно, оказалось, что эпидермальный фактор роста увеличивает число удвоений клеточной популяции с 50 до 150 удвоений с одновременным увеличением "времени жизни" культуры примерно в три раза. При этом увеличение "времени жизни" культуры сопровождалось задержкой проявления многочисленных признаков терминальной дифференцировки кератиноцитов [Rheinwald, Green, 1977].
Другое предсказание гипотезы дифференцировки состоит в том, что, вопреки утверждению Хейфлика, постоянный активный рост клеточных культур может наблюдаться отнюдь не только у трансформированных гетероплоидных клеток, а возможен и для нормальных диплоидных (если эти клетки не вступают на путь терминальной дифференцировки). Действительно, оказалось, что нормальные диплоидные эмбриональные клетки мыши, которые в обычных условиях испытывают кризис роста через 7-10 удвоений популяции, можно успешно культивировать без всяких признаков приближающегося кризиса роста, по крайней мере, в течение 200 удвоений популяции, если только изменить состав культуральной среды (исключить сыворотку крови и добавить ряд компонентов, включая уже упоминавшийся эпидермальный фактор роста). При этом клетки, способные, по-видимому, к неограниченному размножению, остаются диплоидными и незлокачественными [LooetaL, 1987].
Практически неограниченную способность к пролиферации некоторых нормальных диплоидных клеток можно наблюдать не только в культуре (in vitro), но и в условиях целостного организма (in vivo). Этим свойством обладают, например, нормальные клетки имагинальных дисков дрозофилы, если отсутствуют индукторы их дифференцировки [Finch, 1976], а также гемопоэтические стволовые клетки [Harrison, 1984].
С учетом перечисленных выше фактов приходится признать, что концепция Вейсманна-Свима-Хейфлика должна быть пересмотрена, особенно в части тех дополнений, которые были внесены Хейфликом [Hayflick, 1968]. Хотя данная концепция пользуется огромной популярностью, особенно среди исследователей, далеких от клеточной биологии [Fries, 1980; Benjamin, 1982], в настоящее время неясно, какое отношение к продолжительности жизни организмов имеют результаты, полученные при изучении клеточных культур. Не исключено, что ограниченная способность части клеток к делению довольно косвенно связана с ограниченной продолжительностью жизни организмов. В пользу этого вывода свидетельствует также тот факт, что организмы, построенные в основном из постмитотических клеток (лабораторные дрозофилы), имеют принципиально те же закономерности распределения продолжительности жизни, что и человек.
Вместе с тем, очевидно, что переход части клеток в постмитотическое состояние создает предпосылки к снижению регенераторных возможностей организма и в конечном итоге к уменьшению числа функционирующих клеток, что действительно наблюдается с возрастом [Лэмб, 1980; Walton, 1982]. Поэтому исследование кинетики клеточных популяций на разных этапах жизни организма может иметь важное значение для выяснения механизмов, определяющих продолжительность жизни. Однако попытки обосновать существование генетически запрограммированного предела продолжительности жизни ссылками на предел клеточных делений нельзя признать убедительными. Ключ к механизму детерминации продолжительности жизни лежит не столько в выяснении причин снижения темпов пролиферации, сколько в выяснении причин гибели и повреждения клеток, а также других структур организма. Показательно, что один из авторов рассматриваемой концепции предела клеточных делений тем не менее сам отвергает гипотезу генетической программы и отстаивает гипотезу накопления повреждений (износа) [Hayflick, 1988].
См. также:
 5.1. Программа самоликвидации или износ?
5.1. Программа самоликвидации или износ?
5.2. Анализ межвидовых различий по длительности жизни
5.3. Анализ внутривидовых различий по длительности жизни
5.4. Анализ половых различий по срокам жизни
5.5. Эксперименты по продлению жизни
5.7. Теория надежности методологическая основа изучения механизмов, определяющих продолжительность жизни
 Обсудить на форуме
Обсудить на форуме

