С помощью Эск удалось выяснить первые гены, либо комбинации генов, реализующие трехмерную карту зародыша, а также коды линейного созревания клеток-предшественниц в зрелые функциональные единицы органов (дольки печени, альвеолы легкого, нефроны почек). Развитие зародышей с выключенной гамма-субъединицей ламинина LAMC1-/- останавливается на стадии формирования экстраэмбриональной энтодермы из-за дефекта миграции клеток (Smyth N., Vatansever S.H., Murray P. et al., 1999). Нокаут гена Brachyury или zeta-globin вызывал раннюю гибель зародышей из-за блока развития мезодермы. Выключение гена GATA-4 останавливало развитие эндодермы. Вторым master-gene эндодермы оказалась транскриптаза vHNF-1 (variant Hepatocyte Nuclear Factor 1), которая на следующем этапе активировала следующее семейство генов: HNF-4alpha1, HNF-1alpha, HNF-3gamma. Третьим master-gene эндодермы оказался ген cas (casanova) . Выключение этого гена блокировало развитие всей эндодермы зародышей.
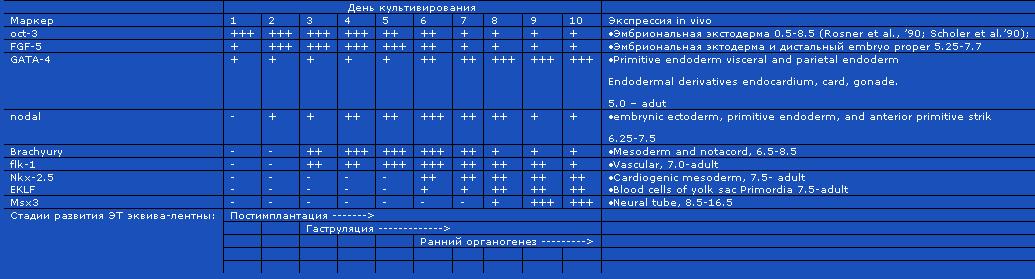
Таблица. Маркеры зародышевых листков в культуре эмбриональных телец
Из табличных данных следует важный вывод: временной порядок включения ключевых транскриптаз совпадает в постимплантационных зародышах и в культуре эмбриоидных телец (вторичных агрегатов Эск) (Leahy A., Xiong J.W., Kuhnert F. Et al., 1999). Порядок включения важнейших генов постимплантационного периода существенно не зависит от факторов микроокружения.
Зародыши мыши GATA3-/- погибали на 11-12 день гестации от блока гемапоэза в фетальной печени. Зародыши мыши SCL-/- погибали на 9.5 день из-за блока желточного кроветворения. Зародыши Flt-/-, как и зародыши Flk-/- погибали вскоре после имплантации из-за дефектов васкулогенеза и капиллярогенеза. Без этих рецепторов ангиобласты подвергались ошибочной сборке. В результате возникали фатальные аномалии в желточном мешке. Если мутация Flk-/- наиболее сильно поражала зародыш (где сосредоточено максимальное число этих рецепторов), то мутация Flt-/- преимущественно блокировала развитие желточного мешка. Если в нокаут-зародыши своевременно пересаживали нормальные Эстволовые клетки, дефект развития устранялся. С помощью донорских клеток удавалось ликвидировать внутриутробную аномалию развития в зародышах-химерах.
Зародыши мыши FGF4-/- останавливались в развитии и погибали сразу после имплантации из-за блока развития клеток трофэктодермы (Nishimoto M., Fukushima A., Okuda A.,1999). Техника нокаут-генов позволила выяснить детали образования транскрипционного комплекса Oct4 / Sox-2. Синергистическая активация/репрессия Oct4/Sox-2 осуществляется с помощью третьего гена -UTF1 (Nishimoto M., Fukushima A., Okuda A.,1999). Показано, что для любых линий Эск in vitro характерна синхронная активация 5 факторов транскрипции: Oct4, Oct6, Sox-2, PEA3 и REX-1. С началом вступления незрелых пролиферирующих Эск в дифференцировку экспрессия 5 факторов транскрипции более или менее синхронно угасает. Экспрессия UTF1 находится под синергестическим контролем генов Oct4/Sox-2. В свою очередь активный белковый комплекс транскриптаз Oct4/Sox-2 активирует третий ген - FGF4. По-видимому, комплекс Oct4/Sox-2 контролирует также экспрессию следующих ранних генов в Эск: osteopontin adhesion molecule (Opn), коактиватор транскрипции UTF1, фактор транскрипции REX-1 (Du Z.,Cong H., Zhen Y.,2001). В ряде линий Эск этот генетический «конвейер» поддержания плюрипотентности клеток нарушается при пассировании. Экспрессия вышеупомянутых факторов транскрипции существенна для оценки качества Эстволовые клетки, поступающих из клеточных банков.
Выключение генов nodal, FGF-5, Oct3 останавливало развитие нейроэктодермы и нервной пластинки. Ген Nkx-2-5, CSX избирательно контролировал закладку кардиогенной мезодермы, Msx3 - закладку нейральной трубки, EKLF - закладку желточного кроветворения (Репин, 2001). Техникой нокаут был идентифицирован SIL ген, ответственный за парное симметричное формирование зачатков органов. На мутантных мышах открыто новое семейство генов Taube Nuss, контролирующих уровень апоптоза клеток у зародышей на стадии органогенеза. Эти гены особенно важны для элиминации провизорных клонов и органов (Voss A.K., Thomas T., Petrou P. et al., 2000). Расшифрованы функции семейства сигнальных белков STAT (signal transducer and activator of transcription) в Эск мышей. STAT1 обеспечивает сигнализацию между рецепторами интерферона и хроматином. Выключение STAT1 элиминировало все клоны зародышей, экспонирующие рецепторы для интерферона. Зародыши STAT4-/- и STAT6-/- развивали летальные дефекты созревания тимоцитов. Зародыши STAT3-/- погибали внутриутробно на стадии раннего органогенеза. Зародыши STAT5a-/- имели внутриутробные аномалии развития молочной железы (Akira S., 1999). У зародышей SF-1-/- нарушалась закладка надпочечников и половых зачатков. У зародышей Wt-1-/- нарушалась закладка и развитие почек. Этим же способом, показано что выключение Нох-1 гена частично блокирует гематопоэз в зародышах. Выключение Нох-7, Нох-8 генов вызывает аномалии развития миоцитов.
После имплантации, клетки эмбриобласта дифференцируются в экстраэмбриональную энтодерму и эмбриональную эктодерму. Эмбриологи искали способы моделирования этих событий in vitro. Если из среды убрать фидер и LIF, пролиферация клонов приостанавливалась. Эск формировали агрегаты (эмбриоидные тельца). В части агрегатов шла дифференцировка клеток, которая в обратном порядке по отношению к зародышу включала маркерные гены трех зародышевых листков. Наружний слой маркировал экстраэмбриональную эндодерму геном GATA-4, тогда как внутренний слой - в эктодерму маркерным геном nodal (Xiong J.W., Battaglino R., Leahy A. et al., 1999). Одновременно в клеточных агрегатах убывала экспрессия гена Oct-4. Несколько лабораторий научились эффективно контролировать дифференцировку клеточных агрегатов (эмбриоидных телец) после экспрессии генов трех зародышевых листков (Schamblott M.J., Axelman J.,Littlefield J.W., et al., 2001). Задача заключалась в направленном получении клеток одного зародышевого листка из эмбриоидных телец. С этой целью научились включать гены гаструляции и органогенеза в эмбриоидных тельцах в более медленном темпе (Leahy A., Xiong J.W., Kuhnert F., J. Exp. Zool., 1999). Включение генов органогенеза автоматически вело к утрате туморигенности постмитотическими клетками эмбриоидных агрегатов. Более того, трансплантационная выживаемость и активная колонизация такой плюрипотентной ткани в организме реципиента была выше, по спавнению с исходными Эск. Пересадки тотипотентной зародышевой ткани на стадии экспрессии первых генов зародышевых листков вытеснят в недалеком будущем пересадки клеточных линий стволовых клеток из-за более высокой эффективности колонизации, приживления, плюрипотентности ростков. По мнению Герхардта, плюрипотентная зародышевая ткань сохраняла синергизм взаимодействия клонов, который необходим для выращивания полноценного трансплантата.Такая плюрипотентная ткань незаменима для моделирования и изучения процессов гаструляции, а также взаимодействия мезенхимы с гомогенной эктодермой или мезодермой in vitro. Напомним, что потенции линий Эск человека в генерации дифференцированных клеток пока не стоит преувеличивать. Например, только 8 % эмбриоидных телец удается in vitro превратить в островки сокращающихся кардиомиоцитов (у Эск мышей эта способность к моно-дифференцировке - на порядок выше) (Kehat I., Kenyagin-Karsenti D., Snir M. Et al., 2001). При этом образующиеся в культуре кардиомиоцитоы имеют незрелую ультраструктцру саркомеров и дисков. Между тем многие биотехнологические компании, владеющие первыми линиями Эск человека, уже делают широковещстельные заявления о том, что линии Эск человека должны стать единственным мостом в будущее ( игнорируя неограниченные геномные возможности природных фетальных тканей).
С помощью Эск апо-Е3-/- удалось вывести линию мышей с высокой предрасположенностью к атерогенному атероскерозу. У этих животных в постнатальном периоде развивалась гиперхолестеринемия и исчезала природная резистентность к стенозирующим бляшкам. Эти мыши сейчас широко используются для тестирования новых гиполипидемических и антиатерогенных препаратов.
Как известно, первичные сигналы, формирующие 3D-оси зародыша после имплантации, возникают на стыке экстраэмбриональной энтодермы, висцеральной энтодермы и эмбриональной эктодермы. Лишь часть этой программы фрагментарно воспроизводится в культуре эмбриоидных телец. (Knezevic V., Mackem S.,2001).
Если Эск выделять из эмбриобласта бластоцисты, первыми возникали эмбриоидные агрегаты с маркерами эндодермы в виде фрагментов желточного мешка. Примитивная висцеральная энтодерма образуется из мигрирующих клеток эмбриобласта. Позднее многие клетки эпибласта мигрируют в париетальную энтодерму и в клетки желточного мешка. Далее из части желточной эндодермы шло формирование первичных ангиобластов и капиллярогенез. Поэтому примесь эндодермы и эндотелиальных клеток удаляли перед повторным пипетированием клонов для получения тотипотентных Эск. Попытки выделить Эск из трофобласта закончились неудачей (возможно из-за наличия в сыворотке бактериальных эндотоксинов). Однако популяцию гигантских клеток трофобласта получают путем трансфекции бластомеров мыши геном Hand1 и его сверхэкспрессии в клетках, которые дифференцируются в трофэктодерму (Fan Y., Melhem M., Chaillet R.,1999). Даже следовые концентрации эндотксина в среде культивирования вызывали массовую гибель незрелых стволовых клеток. Впрочем получить клетки трофобласта из Эск человека или приматов удавалось (Odorico J.S., Kaufman D.S., Thomson J.A., 2001). Недавние исследования израильско-американских биологов выявили комбинации из 8 ростовых факторов, которые направляли дифференцировку эмбриоидных телец в сторону экто-, мезо- или эндодермы. Так, активин в сочетании с TGF-beta предпочтительно стимулировал развитие мезодермы, тогда как комбинация ретиноевой кислоты, bFGF, EGF, BMP-4 стимулировали одновременное развитие экто- и мезодермы. Возникновение эндодермы в культуре не шло без добавления HGF и NGF.
См. также:
 1. На пороге новой биологии и медицины
1. На пороге новой биологии и медицины
2. Эск: основные определения и концепция
3. Основные источники и способы выделения Эск (историческая справка)
4. Молекулярные основы тотипотентности генома Эск
5. Особенности фенотипа Эск
7. Направленная дифференцировка Эск и ППК in vitro
8. Эск в изучении функций Нох-генов
9. Эск - новый биоресурс медицины
10. Эск: законодательство и биоэтика
11. Мост между наукой и клиникой
...
 Обсудить на форуме
Обсудить на форуме

