Д. Бернал писал, что гораздо труднее увидеть проблему, чем найти ее решение. Для первого требуется воображение, а для второго - только умение.
Как уже указывалось, цепные изменения, ведущие к старению и гибели клетки, представляют собой смену накладывающихся друг на друга последовательных этапов - догеномного, генорегуляторного и геноструктурного. Их соотношение неодинаково в различных типах клеток. Важнейшее значение при этом имеют генорегуляторные изменения, ибо именно с ними связан переход от проходящих, компенсируемых, восстанавливаемых сдвигов к стойким, необратимым нарушениям, ибо именно они связаны с возрастными изменениями внутриклеточной и надклеточной регуляции.
Генорегуляторная гипотеза старения была сформулирована нами в 1965 г. (Фролькис, 1965, 1970). В соответствии с этой гипотезой, являющейся составной частью адаптационно-регуляторной теории возрастного развития, важные механизмы старения связаны со сдвигами регуляции генома, саморегуляции генетического аппарата. Это приводит к трем последствиям, которые становятся структурной основой старения клетки: 1) изменению соотношения синтеза различных индивидуальных белков; 2) ограничению надежности, потенциальной возможности биосинтетической системы; 3) раскрытию ранее репрессированных генов, к появлению ранее не синтезированных белков. Эти сдвиги могут происходить как в результате нерепарируемых повреждений генома, так и нарушения ядерно-цитоплазматических, мембраногенетических регуляторных связей.
Регуляторный отдел генома, занимающий большую его часть (до 95% ДНК), чрезвычайно активен в процессе жизнедеятельности, он менее других защищен от внутриклеточных влияний и больше всего подвержен возрастным изменениям. Следует подчеркнуть, что регуляторная часть генома может изменяться, нарушаться, в ней могут возникать апуриновые сайты, разрывы, димеры, делеции, ошибки, которые приводят к изменению регулирования генома. Информация, заложенная в ДНК, может закономерно реализовываться благодаря существованию структурно закрепленной системы - хроматина. Это и определило большое внимание геронтологов к этому механизму реализации генетической информации. Так, показано, что при старении происходят хромосомные аберрации, увеличение содержания гетерохроматина, увеличение перекрестных связей между ДНК и белками, дисульфидных связей белков, а также снижение матричной активности ДНК, отношения РНК/ДНК и др. (Chaturvedy, Kanungo, 1983).
В нашем коллективе И. Ю. Хилобок и T. H. Мозжухиной (1983) было проведено изучение возрастных особенностей состава, структурной организации и матричной активности активной и неактивной фракций хроматина печени крыс.
Оказалось, что при старении происходит перераспределение относительного содержания фракций хроматина, рассчитываемых в процентах от суммарной ДНК хроматина. У старых животных достоверно увеличивается доля неактивной фракции и соответственно уменьшается содержание активной. В неактивной фракции хроматина печени не изменяется содержание РНК и белка, в активной отношение белок/ДНК увеличивается на 20 %, более прочной становится связь белков с ДНК (рис. 2).
При электрофорезе фракций хроматина печени, представленных набором нуклеосом и кратных им фрагментов, образующихся при фрагментации хроматина Ca-, Mg-зависимой эндонуклеазой, было установлено, что препараты хроматина старых животных содержат меньше мононуклеосом и больше олигомеров нуклеосом.
В неактивной фракции хроматина печени относительное количество мономерных фрагментов составляет 30 и 17 %, в активной - 16 и 12 %; количество олигомеров размером более 4 нуклеосом в неактивной фракции 29 и 41 %, в активной - 33 и 41 % соответственно для взрослых и старых крыс. В то же время исследование скорости выхода суммарных фрагментов в условиях активации эндонуклеазы не выявило возрастных различий. Полученные результаты, таким образом, свидетельствуют об изменении при старении процесса фрагментации хроматина печени, связанного с изменением доступности участков расщепления хроматина к действию Ca-, Mg-эндонуклеазы, вследствие уплотнения его структуры на наднуклеосомном уровне.
При сопоставлении электрофоретической подвижности фрагментов хроматина было обнаружено снижение подвижности нуклеосом и кратных им фрагментов неактивной фракции хроматина печени старых крыс по сравнению со взрослыми. Это снижение обусловлено увеличением размера нуклеосомной ДНК, выщепляемой эндонуклеазой, что подтверждается результатами электрофореза ДНК, выделенной из соответствующих фракций хроматина печени взрослых и старых крыс.
Итак, при старении происходит не только наднуклеосомная компактизация хроматина, но и изменение доступности линкерной ДНК к действию фермента.
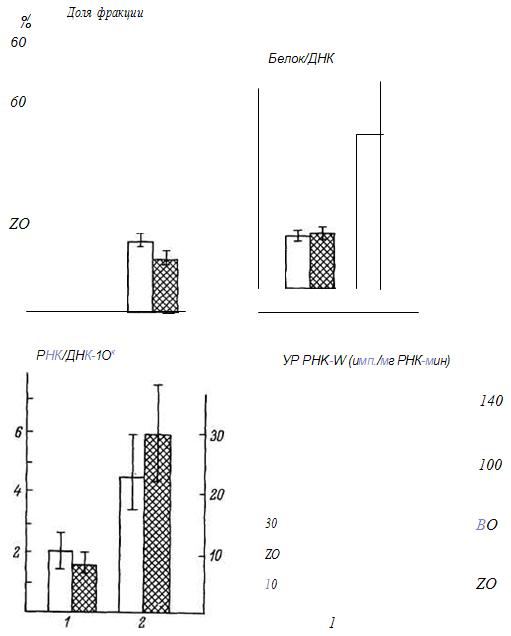
Рис. 2. Характеристики неактивной (/) и активной (2) фракций хроматина печени крыс разного возраста.
Светлые столбики - взрослые; заштрихованные - старые.
Возрастная компактизация хроматина обусловлена изменением взаимодействия ДНК и белков хроматина, упрочнением их связи, о чем свидетельствует увеличение температуры плавления фракций хроматина печени. Следует отметить, что увеличение температуры плавления при старении более выражено в активной фракции хроматина. В. И. Никитин (1962) полагает, что накопление в хроматине прочных связей ДНК-белок является одним из основных пусковых механизмов старения.
Таким образом, результаты изучения процесса термоденатурации, так же как и данные электрофоретического анализа фракций, указывают на увеличение плотности упаковки хроматина печени при старении за счет дополнительной стабилизации участков свободной и линкерной ДНК и на увеличение относительной доли плотно упакованных структур. Эти изменения, по-видимому, лежат в основе перераспределения фракций активный-неактивный хроматин и в снижении транскрипционной активности хроматина печени. Интересно предположение M. M. Виленчика (1978): благодаря компактизации снижается доступность поврежденных участков ДНК для ферментов репарации. Слабость репарации сама по себе способствует увеличению числа сшивок ДНК и белков. Весь этот комплекс изменений хроматина может стать основой возрастных изменений регулирования генома и последующих сдвигов в процессе биосинтеза белка.
Обращает на себя внимание то, что описанные изменения хроматина возникают у старых животных в возрасте 28-30 мес, а у 24-26-месячных их нет. Очевидно, это следствие возникших ранее менее глубоких сдвигов. Существует еще ряд доказательств изменения регулирования генома в процессе старения.
1. Изменяется спектр, соотношение синтезируемых РНК. Используя метод гибридизации, Гаубатц, Катлер (Gaubatz, Cutler, 1978), Стрелер и Чанг (Strehler, Chang, 1979) показали, что в мозге, печени, почках, селезенке-старых людей на 9-40 % уменьшается число копий ДНК, ответственных за синтез рибосомальной РНК. Это нарушение регуляции генома, очевидно, связано с накоплением у старых животных сшивок между ДНК и белками. Действительно, интенсивная депротеинизация однонитевой ДНК приводила к восстановлению уровня гибридизации у старых животных. Катлер (Cutler, 1979) показал, что при старении число разновидностей вновь синтезированной РНК снижается.
По нашим данным, изменяется соотношение скорости синтеза различных классов РНК. Так, если относительную УР прерибосомальной РНК (пре-рРНК) ядра гепатоцитов принять за единицу, то соотношение УР пре-рРНК, смеси пре-рРНК и преинформационной РНК (пре-иРНК), пре-иРНК и гетерогенной ядерной РНК у новорожденных крысят составит 1:1:1:1, у одномесячных - 1:2:1:1, у 6-8-месячных - 1:3:2:2, у 24-месячных - 1:2: 1 : 1, у 38-месячных - 1:3:4:2. Кроме того, с возрастом изменяется соотношение специфических форм РНК внутри одного класса, например, соотношение транспортных РНК, переносящих различные аминокислоты.
2. Изменяется соотношение синтеза отдельных белков. Имеется немало данных, свидетельствующих о неравномерных сдвигах синтеза белков крови, печени, миокарда. Изменяется изоферментный спектр многих ферментов. В нашем коллективе показано, что при старении изменяется соотношение изоформ ЛДГ, креатинфосфокиназы. Так, у старых животных увеличивается активность ЛДГ в скелетной и сердечной мышце и не изменяется в печени. При этом в скелетной мышце увеличено содержание М-субъединиц, а в сердечной - Н-субъединиц (Литошенко, 1984).
Известно, что существует три изоформы миозина, отличающиеся по АТФазной активности, пептидным картам, антигенным свойствам. При старении меняется соотношение изоформ миозина - растет количество формы V3 и падает У, (Syrovy, 1984). Это и приводит к снижению активности Ca2+ АТФазы миозина в желудочках сердца в старости. Сдвиги регулирования генома могут быть столь существенны, что вызывают изменения соотношения различных полипептидных цепей в белковой молекуле. Так, в старости нарастает содержание фетального гемоглобина, состоящего из двух а- и у-полипептидных цепей (а2, у2) в отличие от гемоглобина зрелых животных - типы а2, |32.
3. Известна роль гистоновых и негистоновых белков в регуляции генома. Гистоны в большей мере оказывают репрессирующее влияние, а негистоновые белки - дерепрессирующее. Учитывая многочисленность этих белков, необходимо, конечно, с осторожностью использовать подобное разделение эффектов. С возрастом соотношение негистоновые белки-гистоны существенно изменяется. В печени 3-месячных крыс оно равно 0.62±0.01; 12-месячных - 0.53±0.01; 24-месячных - 0.43±0.01. Предполагается, что это изменение соотношения белков влияет на регуляцию генома в старости.
При старении изменяется метилирование, фосфорилирование, ацетилирование белков хроматина (Ванюшин, 1983; Клименко, 1984). Так, заметно снижается фосфорилирование богатых лизином гистона H1 и кислых белков, глобулиновых белков, резко падает ацетилирование лизинбогатых гистонов и увеличивается ацетилирование кислых белков. Эти модификации белков изменяют подвижность их связи с ДНК и могут стать причиной сдвигов регуляции процесса транскрипции определенных генов, приводить к увеличению компактизации хроматина. Существенное значение в регуляции активности генома имеет метилирование ДНК- Определенные последовательности, выполняющие функции стартовых точек для считывания сцепленных генов, вероятно, являются одновременно специфическими местами метилирования. Б. Ф. Ванюшин (1982) подчеркивает, что возрастные изменения уровня метилирования ДНК затрагивают главным образом регуляторные, а не структурные гены. Так, содержание одного из минорных соединений - 5-метил-цитозина - может к старости падать в 1.5-2 раза.
4. Обоснованно предположение, что значительные изменения регуляции генома при старении могут привести к активации ранее молчавших генов, к синтезу ранее несинтезированных белков. В зависимости от его типа могут возникнуть различные нарушения деятельности клеток вплоть до их гибели. Однако это предположение нуждается в конкретных экспериментальных доказательствах.
5. Важный молекулярный адаптационно-регуляторный феномен, проявление процесса витаукта - генетическая индукция ферментов, определяемая регулированием генома. Благодаря синтезу ферментов клетка адаптируется к изменившимся условиям существования, измененному уровню деятельности. В нашем Институте были показаны возрастные изменения генетической индукции ряда ферментов переаминирования аминокислот, глюконеогенеза, микросомального окисления в печени (Мурадян, 1976; Парамонова, 1981). Многие противоречия в описании изменения генетической индукции в старости связаны с тем, что при этом не характеризуется реактивность соответствующей системы. Речь идет об использовании широкого диапазона доз субстратов и гормонов. Оказалось, что при малых дозах гидрокортизона и тирозина сдвиги в индукции тирозинаминотрансферазы более выражены у старых животных, при средних возрастные различия нивелируются, а при больших дозах сдвиги активности больше у взрослых животных. Подобные закономерности отмечены при изучении индукции триптофанпирролазы, глюкозо-6-фосфатазы; фруктозо-1,6-дифосфатазы. Интенсивность индукции ферментов микросомального окисления с возрастом падает. Следует указать, что подобный тип изменения генетической индукции не является универсальным для всех ферментов.
6. Генорегуляторные возможности в старости ограничены. Для характеристики системы биосинтеза белка, как и для любой системы организма, важно оценить ее возможности, надежность в условиях напряженной деятельности, при даче нагрузок. По данным С. H. Новиковой (1978), после кровопотери у старых животных возникает менее выраженная активация биосинтеза белка в печени, медленнее восстанавливаются белки крови. По данным Ф. 3. Меерсона (1981), при гиперфункции миокарда у старых крыс в меньшей мере активируется синтез РНК и белков. Существенная активация синтеза рДНК, иРНК белка возникает при регенерации после гепатэктомии. По данным E. Л. Левицкого (1984), выраженность этой активации с возрастом падает. Таким образом, ограничение потенциальных возможностей биосинтетических систем выявляется в различных условиях напряженной деятельности в старости.
7. В механизме возрастных изменений биосинтеза белка существенное значение имеют сдвиги в ионном гомеостазе (Фролькис, 1975). На разных объектах показано существование зависимости характера белкового синтеза от соотношения ионов Na+H K+ (Данко и др., 1984). Одновалентные ионы оказывают влияние как на процессы транскрипции, так и трансляции, на системы ферментативного контроля за реализацией транскрипционной программы. В клетках одного типа биосинтез белка активируется при увеличении концентрации K+, другого - Na+. Более того, некоторые гормоны оказывают влияние на последовательность синтеза определенных белков опосредованно через ионные сдвиги в клетке.
В процессе старения изменяется ионный гомеостаз в клетках различного гистогенеза. По данным одних авторов, в ряде клеток нарастает концентрация Na+ (Тураева, 1978), по данным других - K+. Нами прямо показано, что при изменении ионного состава клетки, вызванном гиперполяризацией, активируются процессы транскрипции и трансляции и эта активация слабее выражена у старых животных.
Через ионные сдвиги осуществляются мембранные механизмы регуляции синтеза белка, связь между функцией клетки и активностью генов. Изменение этих связей - важный механизм старения клетки.
Большинство исследователей приходит к выводу о снижении скорости синтеза суммарного белка, которое и становится ведущим механизмом старения клеток. По нашим данным, изменение синтеза белков неодинаково выражено в различных органах. Канунго (Kanungo, 1982) полагает, что органная специфичность связана с неодинаковой ролью этих белков, изоферментов в различных клетках. На фоне снижения синтеза суммарных белков синтез отдельных белковых молекул может изменяться неодинаково. Так, например, в гепатоцитах снижается синтез ферритина, но нарастает синтез альбуминов.
Существенное значение в изменении синтеза белков имеют сдвиги в трансляции - системе аминоацилирования тРНК, рибосомного и полисомного аппарата, белковых факторов трансляции, эндогенных ингибиторов биосинтеза белка и др. (Бердышев, Никитин, 1982). Таким образом, при старении наступают изменения во всех звеньях прямой связи биосинтеза белка - на этапе транскрипции и трансляции. Значение генорегуляторных изменений состоит в том, что они наиболее стойко изменяют синтез определенных белков.
Существует два уровня регуляции белковых молекул, активности ферментов - аллостерическая регуляция (изменение конформации белковых молекул) и генетическая индукция (образование белков de novo). При старении изменяются оба типа регуляции - не только ограничивается диапазон генетической индукции ряда ферментов, но и во многих случаях под влиянием изменения функции клетки, физиологически активных веществ в меньшей мере растет активность ферментов.
Как известно, белковые молекулы стареют. Период полужизни различных белков колеблется в больших пределах - от нескольких минут до периодов, равных жизни организма (коллаген). Чем больше ВПЖ, тем более выражены возрастные изменения структуры коллагена. И. M. Спектор (1974) обнаружил прогрессивное увеличение периода полужизни меченных иодом альбуминов и глобулинов по мере увеличения ВПЖ.
Предполагается, когда ферментативные белки становятся старыми, они элиминируются из организма. В основе старения белковых молекул лежат два типа изменения: 1) накопление посттрансляционных ошибок в структуре белка. Клоповская и Клоповский (Kjopowska, Klopowsky, 1985) предполагают, что чем больше молекула белка, тем больше в нем возникает ошибок и тем быстрее он стареет, 2) переход белка из нестабильного (активного) в стабильное (неактивное) состояние благодаря изменениям в его третичной и четвертичной структуре. Вероятно, чем больше молекулярная масса белка, тем медленнее наступает старение этого типа.
Изучение временной нестабильности структуры и регуляции генома получило новый импульс после открытия мобильных диспергированных генов (МДГ). Анализ этих работ совместно с данными о возрастных сдвигах генома и смертности убеждает нас в целесообразности рассмотрения еще одной гипотезы старения - гипотезы «горячих точек».
Одним из крупных открытий молекулярной биологии стало доказательство мозаичной структуры генов. Оказалось, что участки, кодирующие белки (экзоны), разорваны некодирующими фрагментами (интроны). Сперва образуется РНК, транскрибируемая как с экзонов, так и с интронов. Затем в процессе сплайсинга участки РНК, транскрибируемые с интронов, вырезаются, а с экзонов - соединяются. Экзоны соответствуют функциональным фрагментам белковых молекул и за счет интронов может происходить перекомбинирование экзонов в эволюции, приводящее к образованию новых белков. Предполагается, что фрагменты РНК, считанные с интронов, могут регулировать экспрессию генов по принципу обратной связи.
Генорегуляторные изменения при старении могут быть связаны с неизбежно повторяющимся сплайсингом. Фрагменты РНК могут изменять экспрессию генов, накопление их, могут влиять на состояние хроматина. Интересно, что ингибирование процессов транскрипции увеличивает продолжительность жизни, а стимуляция ее - сокращает.
Современная молекулярная биология исходит при объяснении первичных механизмов старения из принципа «один ген - один белок», и старение клетки при этом рассматривается как результат последовательного нарушения регуляции синтеза белков. Однако есть генетическая информация, недостаточно еще изученная, определяющая общую структуру клетки, ее метаболизм, число клеток в органе, межклеточные отношения, размер, форму всего органа и др. Можно полагать, что наиболее общие механизмы старения клетки и связаны с нарушением этого уровня регуляции. Мы назвали его алгоритмическим. Именно описываемый генетический механизм объясняет системность нарушения в процессе старения. Именно благодаря ему можно понять связь между старением и витауктом. В соответствии с ним страдает алгоритм, «программа», порядок включения «генетической информации» в клетке, что приводит к бессвязности текста, разрегулированию обмена и функции клетки как целостной структуры. Именно этим можно объяснить множественность проявлений старения клетки, нарушение ее функции вследствие неглубоких изменений в отдельных звеньях системы. При таком системном подходе нарушение синтеза отдельного белка - только звено в возрастном изменении генопрограммы. На основании алгоритмического механизма объяснимы отличия в старении клеток, имеющих различные функции.
Следует разграничивать внутриклеточные и надклеточные нейрогуморальные механизмы саморегуляции, биосинтеза белка. Большую роль в поддержании гомеостазиса клетки играют ядерно-цитоплазматические отношения. По данным Деви и др. (Devi et al., 1966), при добавлении к ДНК молодых крыс цитозола из печени старых уровень синтеза РНК оставался таким же высоким, как у молодых. При добавлении к ДНК старых животных цитозола гепатоцитов молодых синтез РНК по-прежнему оставался на низком уровне. Билка и сотр. (Bielka et al., 1976) показали, что очищенные рибосомы печени и почек крыс в бесклеточной среде с полиуридином и Н3-фен-тРНК не выявляли возрастной разницы в скорости синтеза белка. Вместе с тем активация этого синтеза цитозолом почек и семенников резко падала с возрастом. По данным Гельтхаллера и др. (HeIIthaler et al., 1976), в старости существуют иные ядерно-цитоплазматические отношения. Добавление цитозола из печени старых крыс к микросомам 3-5-месячных крыс приводило к снижению их белоксинтезирующей способности до уровня старых крыс, а добавление цитозола печени 3-5-месячных животных к микросомам старых вызывало повышение интенсивности синтеза белка до уровня молодых. Нами и H. С. Верхратским было показано, что добавление цитозола ткани печени старых крыс подавляло синтез информационной и рибосомальной РНК в ядрах молодых животных. Цитоплазма активизировала синтез в старых ядрах, хотя уровень синтетических процессов оставался ниже, чем в другие возрастные периоды.
Удачными для выяснения возрастных изменений регуляторных связей оказались опыты с реконструированием клеток. Магглетон-Харрис и Хейфлик (Muggleton-Harris, Hayflick, 1977) реконструировали клетки W-138 из цито- и кариопластов молодых и старых клеток и исследовали образованные ими клоны. Оказалось, что клетки, реконструированные из старых кариопластов и молодых цитопластов, погибали через четыре недели после слияния и не были способны к репликации. Клетки, реконструированные из молодых кариопластов и старых цитопластов, живут 16 недель и делятся 3 и более раз. Это, однако, меньше, чем у клеток, образованных из молодых карио- и цитопластов. Подводя итоги обзору работ по исследованию старения в опытах с клетками опухолей и с клетками, трансформированными in vitro, A. В. Зеленин и др. (1982) полагают, что гибридным клеткам не удается преодолеть старение в фибробластах, однако некоторое омоложение в гибридах происходит. Интересны данные МагглетонаХарриса и Пезелла (Muggleton-Harris, Pezella, 1979), помещавших ядра старых эпителиальных клеток хрусталика в энуклеированные яйцеклетки лягушек того же вида. Определенное число возникших зародышей развивалось в нормальные лягушки. Банн и Торрент (Bunn, Torrant, 1980) показали омолаживающее действие цитоплазматических клеток на старые фибробласты. В результате образования таких гибридов многие ядра старых клеток приобрели способность вступать в S-фазу и давать около 20 делений.
Все эти опыты убеждают в том, что геномные изменения при старении не являются необратимыми, а в определенных ситуациях клетка может «омолаживаться». Они подтверждают и представление о стохастичности развития старения, во многом формирующемся в ядре под влиянием цитоплазматических факторов, об участии как ядра, так и цитоплазмы клетки в развитии ее старения.
Таким образом, при старении изменяются как регуляторные воздействия, идущие с цитоплазмы, так и состояние самого ядерного синтеза РНК. Влияния цитоплазмы на ядро столь существенны, что во многих клетках в старости возникает многоядерность, - полиплоидия - выражение процесса витаукта.
Сложные взаимоотношения возникают в клетке между ядерным геномом и митохондриями. Как известно, митохондрии обладают своим геномом, своей системой синтеза белка. Так, показано, что около 10 % белков митохондрий кодируется митохондриальным геномом, а остальные - ядерным. Митохондриальный геном кодирует ряд субъединиц ферментов дыхательной цепи и протонной АТФазы. А. Я. Литошенко (1985) установил некоторые возрастные особенности митохондриального генома. Оказалось, что в печени старых крыс снижена скорость синтеза митохондриальной ДНК (на 40 %), хотя активность митохондриальной полимеразы, синтезируемой под контролем ядерного генома, не изменяется. По данным А. Я. Литошенко, причиной снижения скорости синтеза митохондриальной ДНК является замедление процесса репликации. В митохондриях гепатоцитов и кардиомиоцитов изменяется соотношение белков, кодируемых ядерным и митохондриальным геномом.
Мы полагаем, что в старении клетки существенное значение имеют открытые нами мембранно-генетические механизмы. Нами было обнаружено, что при активации биосинтеза белка развивается гиперполяризация клеточной мембраны (Фролькис, 1978, 1981, 1982). Это было показано на примере различных клеток (гепатоциты, нейроны, скелетно-мышечные клетки, клетки эндокринных желез) при действии самых различных факторов (гормоны-инсулин, глюкортикоиды, половые стероиды и др.; кровепотеря, регенерация и др.). Во всех этих случаях гиперполяризация сопровождалась активацией биосинтеза белка (актиномицин D, оливомицин, пуромицин, циклогексимид предупреждали развитие гиперполяризации клеточной мембраны). При активации биосинтеза белка в клетке синтезируется специальный гиперполяризующий фактор пептидной природы (Фролькис, 1982).
Оказалось, что возникающая гиперполяризация плазматической мембраны по принципу обратной связи влияет на активность генетического аппарата, на биосинтез белка. В большинстве случаев эта обратная связь подавляет биосинтез белка. В гепатоцитах старых животных угнетающее влияние более выражено, чем у животных взрослых (рис. 3). Активация биосинтеза белка в печени крыс вызывалась гидрокортизоном или фенобарбиталом. На фоне этого анодом постоянного тока добавочно воспроизводилась гиперполяризация. Как видно на рис. 3, у старых животных анодная гиперполяризация в большей мере подавляла генетическую индукцию ферментов, синтез РНК и белков.
Этот эффект гиперполяризации не может быть сведен к снижению входа в клетку индукторов, так как прямое определение показало, что при этом не снижается количество их в клетках. Итак, в клетке существует важный механизм саморегуляции биосинтеза белка - активация генома ведет к гиперполяризации клеточной мембраны, а гиперполяризация по принципу обратной связи угнетает биосинтез белка.
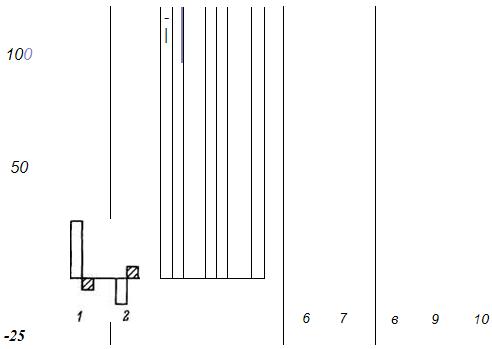
Рис. 3. Влияние гиперполяризации плазматической мембраны на индуктивный синтез ферментов и интенсивность синтеза РНК и белка в печени взрослых и старых крыс.
1 - цитохром Р-450; 2 - аминопириндеметилаза; 3 - фруктозо-1,6-дифосфатаза; 4 тирозинаминотрансфераза; 5 - триптофан-пирролаза; 6 - РНК", 7 - белок; 8 - хроматин активный; 9 - неактивный; JO - гомогенат. Светлые столбики - взрослые; заштрихованные - старые.
Нами, Г. И. Парамоновой и H. Б. Гольдштейн Гольдштейн было изучено влияние первичной анодной гиперполяризации гепатоцитов на распределение хроматина, его транскрибирующую способность, биосинтез белка в клетках печени. Оказалось, что в этом случае гиперполяризация активирует геном и биосинтез белка (рис. 3). Важно то, что это активирующее влияние более выражено у взрослых животных. Следовательно, гиперполяризация в зависимости от исходного состояния генома может оказывать двоякое действие на биосинтез белка - подавляет или стимулирует его. У старых животных в большей мере, чем у взрослых, выражено подавляющее действие и в меньшей стимулирующее. Все это - важный фактор изменения клеточной регуляции биосинтеза белка в старости. Итак, в развитии старения существенное значение имеет изменение внутриклеточных механизмов саморегуляции генома как на уровне самой структуры генома - хроматина, так и ядерно-цитоплазматических, ядерно-митохондриальных, мембранно-генетических отношений. В конечном итоге именно это и определяет ограничение приспособительных возможностей биосинтетических систем в старости, ограничение пластического обеспечения функции.
Существенные сдвиги в процессе старения возникают в надклеточной, нейрогуморальной регуляции биосинтеза белка. Особенное значение имеет изучение возрастных изменений гипоталамической регуляции биосинтеза белка. Во-первых, гипоталамические структуры через гормональные и нервные пути регулируют состояние внутренней среды организма; во-вторых, в естественных условиях при осуществлении ряда важнейших адаптивных реакций регуляция обменных процессов в клетках осуществляется через гипоталамические структуры; в-третьих, многие исследователи полагают, что с нарушением функции гипоталамуса связаны важные механизмы старения на уровне целостного организма.
В цикле наших работ (Фролькис и др., 1971, 1976, 1979) было показано, что при стимуляции структур гипоталамуса у старых крыс возникает менее выраженная, а порой и качественно другая, чем у взрослых, генетическая индукция глюкозоб-фосфатазы, фруктозо-1,6-дифосфатазы, а также активация синтеза РНК. Так, после однократной стимуляции вентромедиального ядра гипоталамуса активность глюкозо-6-фосфатазы росла у зрелых крыс на 195 %, фруктозо-1,6-дифосфатазы - на 186 %, а у старых животных падала на 34 и 17 % соответственно. При многократном раздражении гипоталамуса у старых крыс развивается подавление синтеза фракций РНК и активности ферментов. Было показано, что в этом случае деятельность всей регулируемой системы гипоталамус-гипофиз-кора надпочечников-печень лимитировалась сдвигами, происходящими в самом гипоталамусе. Важно было проанализировать особенности взаимодействия двух важнейших регуляторных систем - на уровне клетки - хроматин, на уровне целостного организма - гипоталамус. Это было проделано нами и И. Ю. Хилобок, Ю. E. Рушкевичем, H. Б. Гольдштейн. Само введение электрода в область вентро- 3 В В Фролькис, X К Мурадян медиального ядра гипоталамуса изменяет его функциональное состояние. В старости гипоталамус становится менее надежной системой, и в результате этого само введение электрода более выраженно изменяет его состояние, о чем свидетельствует неизменность в этих условиях хроматина гепатоцитов взрослых крыс и сдвиги в его состоянии у старых. Электрическая стимуляция у старых животных в меньшей степени влияет на состояние хроматина, биосинтез белка.
Итак, стимуляция гипоталамуса влияет на состояние хроматина гепатоцитов, и этот контроль с возрастом существенно изменяется. Влияние гипоталамуса на биосинтез белка в печени - одно из звеньев общего адаптационного синдрома, сопровождающегося активацией глюконеогенеза, системы микросомального окисления.
Таким образом, при старении существенно изменяется внутриклеточная и надклеточная регуляция генома клетки. Признание генорегуляторного механизма старения обосновывает реальность попыток нормализации деятельности генома, возможность использования этого пути с целью увеличения продолжительности жизни.
См. также:  Функция клеток.
Функция клеток.
 Обсудить на форуме
Обсудить на форуме

