Нет, бытие - не зыбкая загадка;
Подлунный дом и ясен, и росист.
Мы - гусеницы ангелов; и сладко
Въедаться с краю в нежный лист.
Владимир Набоков
Плодовая мушка Drosophila melanogaster - также хорошая модель для изучения генетических компонентов долголетия. Как и в случае с С. elegans, мутанты дрозофилы, имеющие разную продолжительность жизни, были индуцированы с помощью химического мутагенеза. Среди полученных линий некоторые имели мутации в гене Cu,Zn-супероксиддисмутазы (СОД). Гомозиготные по этому гену мутанты развивались нормально, но продолжительность их взрослой жизни сократилась с 60 до 10 дней (Phillips et al., 1989). Было показано, что эти мутанты имеют повышенную чувствительность к веществам, продуцирующим свободные радикалы, и имеют недостаточно активную сперму, что может указывать на важность СОД в защите ДНК от повреждений в ходе гаметогенеза. С другой стороны, особи с увеличенным числом копий генов СОД и каталазы имели большую на 20-37 % среднюю и максимальную продолжительность жизни, тогда как мухи с избыточными копиями генов лишь одного из этих ферментов антиокислительной защиты таким эффектом не обладали (Orr, Sohal, 1994). Трансгенные дрозофилы с избыточной экспрессией гена SOD1 в мотонейронах жили на 40% дольше и были значительно устойчивее к окислительному стрессу, чем мухи, не имевшие этого гена (Parkes et al., 1998). Использовав специальную методику получения трансгенных дрозофил с геном Cu,Zn-SOD, сверхэкспрессированным короткими нагреваниями, активирующими регулируемую промотором hsp70 рекомбиназу FLP, удалось получить линию с увеличенной на 48 % продолжительностью жизни (Sun, Tower, 1999). Избирательная сверхэкспрессия одного гена каталазы в этих опытах не сопровождалась увеличением продолжительности жизни, а когда избыточно экспрессировались гены как СОД, так и каталазы, мухи жили меньше, чем только с трансгеном Cu,Zn-SOD.
При инсерции гена белка теплового шока Hps70 установлена положительная корреляция между числом копий гена (количеством экспрессированного белка) и продолжительностью жизни дрозофил (Tatar et al., 1997). Эти наблюдения, как и ряд других, свидетельствуют о важной роли белков теплового шока Hsp для долголетия. Было также установлено, что сверэкспрессия Hsp70 может защищать нейроны от дегенерации в дрозофильных моделях нейродегенеративных заболеваний, например болезни Паркинсона или Хантигтона (Link, 2001).
У дрозофил система передачи сигнала от инсулина и инсулинподобного фактора роста 1. IGF-1 регулирует размеры тела и так же, как у нематод, продолжительность жизни. Мутации в регуляторном участке гомолога рецептора инсулина (chico) у дрозофил сопровождаются увеличением продолжительности жизни на 45 % (Clancy et al., 2001). Если же мушки были гетерозиготны по двум различным мутациям в рецепторе инсулина/IGF-1, то продолжительность их жизни была на 85 % дольше дикого типа (Tatar et al., 2001). При этом мушки были стерильны, и их яичники напоминали яичники мух дикого типа в диапаузе. Оба типа мутаций в рецепторе инсулина сопровождались также увеличением активности СОД. Хотя параллели между системами регуляции инсулина у нематод и плодовых мух весьма отчетливы, имеются и определенные различия. Полагают, что не все компоненты механизма передачи сигнала инсулина имеют отношение к регуляции продолжительности жизни (Kenyon, 2001).
Lin и соавт. (1998) идентифицировали линию дрозофил с инсерцией Р-элемента транспозона, которые жили на 35% дольше особей дикого типа. Эта мутация, названная mth ("мафусаилова" от Methuselach - Мафусаил), повышала устойчивость к стрессу, вызванному самыми различными факторами, такими как голодание, нагревание или добавление в корм генератора активных форм кислорода параквата.
Среди линий плодовых мух с инсерцией P-элемента были обнаружены долгоживущие мутанты Indy (I am not dead yet) с двукратно увеличенной средней продолжительностью жизни и увеличенной на 50% максимальной продолжительностью жизни (Rogina et al., 2000). Этот ген кодирует белок, гомологичный ко-переносчику Na-дикарбоксилазы млекопитающих, ответственной за захват и обратный захват таких субстратов в цикле Кребса, как янтарная кислота, цитрат и альфа-кетоглютарат.
Также к увеличению продолжительности жизни дрозофил приводит мутация в рецепторе стероидного гормона экдизона. (Simon et al., 2003). Гетерозиготные мухи с такими мутациями живут на 40-50% дольше мух дикого типа и характеризуются повышенной устойчивостью к стрессу. У них отсутствуют дефекты в овогенезе и сперматогенезе, что позволяет предполагать, что увеличение продолжительности жизни у этих мутантов обусловлено изменениями в репродуктивной системе. У гетерозиготных EcR мух фертильность и плодовитость увеличивалась по сравнению с диким типом. Таким образом, увеличение продолжительности жизни мух снижением активности рецепторов к экдизону (EcR) не приводит к утрате или снижению репродуктивного потенциала. Следует отметить, что увеличение продолжительности жизни наблюдали при мутациях в рецепторе экдизона у дрозофил другой линии (Canton-S), а также при индукции мутаций в других локусах, в частности в домене связывающего лиганда и ДНК связывающего домена, и при реципрокном скрещивании мутантов (Simon et al., 2003). Авторы этой работы полагают, что по крайней мере два различных компонента одного метаболического пути (рецептор и лиганд) могут контролировать долголетие у мух. Интересным и важным является наблюдение, что мутации, приводящие к увеличению продолжительности жизни, увеличивают также фертильность и плодовитость дрозофил.
Ускоренное старение у D. melanogaster является результатом не только мутаций в специфических генах, но может также быть результатом изменений в экспрессии эпигенетических факторов. Так, у старых мушек было обнаружено резкое снижение экспрессии белка фактора элонгации EF-1a, предшествующее общему снижению синтеза белков. Когда с помощью генетических манипуляций в геном мух были добавлены дополнительные копии гена EF-1a (Shepherd et al., 1989), то наблюдали значительное увеличение продолжительности жизни этих мух.
Таблица 5. Мутации, увеличивающие продолжительность жшпп у D. melanogaster (Aigakl et aL, 2002, с модификациями)
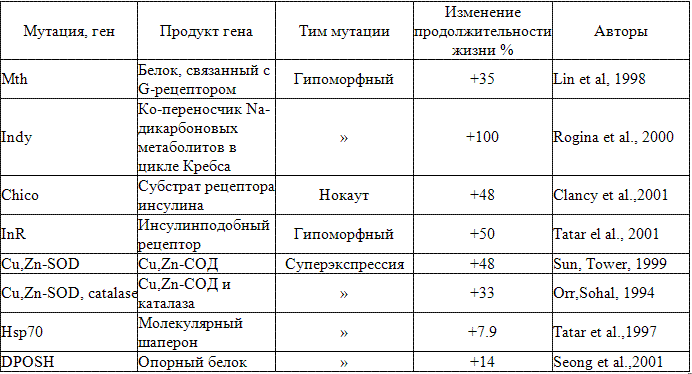
Суммируя результаты исследований по мутагенезу у дрозофил, приводящему к увеличению продолжительности их жизни (табл. 5), Aigaki и соавт. (2002) подчеркивают, что такие мутации не могут быть частыми. Важно не только, какой ген мутирован, но и как он мутирован. Другое потенциальное ограничение заключается в плейотрогшости мутации.
См. также:
 1.1. Популяционная генетика старения
1.1. Популяционная генетика старения
1.2. Наследственное преждевременное старение
1.3. Репродуктивное поведение и эволюция продолжительности жизни
1.4. Гены гибели и долголетия у круглых червей
1.6. Гены долголетия у мышей
1.7. Предполагаемые гены смерти и долголетия человека
1.8. Роль специфических хромосом в старении
1.9. Изучение профиля генетического транскриптома при старении
1.10. Заключение
 Обсудить на форуме
Обсудить на форуме

