Чтоб мудро жизнь прожить, знать надобно немало.
Два важных правила запомни для начала:
Ты лучше голодай, чем что попало ешь,
И лучше спи один, чем вместе с кем попало.
Омар Хайям
Один из ключевых вопросов геронтологии заключается в выяснении роли генетических факторов в старении. Другими словами, определяется ли генами продолжительность жизни животных и человека? Казалось бы, существенные различия в видовой продолжительности животных различных видов однозначно положительно решают этот вопрос. Так, эти различия варьируют до 1 миллиона раз среди всех видов, живущих на Земле и от 10 до 50 раз внутри групп с одинаковым уровнем организации (Carey, Judge, 2000). Среди млекопитающих рекорд долгожительства принадлежит одной из пород китов, продолжительность жизни которых превышает 200 лет. Вместе с тем лабораторные грызуны живут не более 2-3 лет, тогда как многие другие грызуны такого же размера живут до 5-10 лет и более (Finch, Ruvkun, 2001). Большая вариабельность наследования продолжительности жизни организмов одного вида (табл. 2) также не может быть интерпретирована однозначно (Finch, Ruvkun, 2001).
Проанализировав 28 таблиц дожития линий D. melanogaster, контрастных по продолжительности жизни и ряду физиологических характеристик, а также их гибридов, С. В. Мыльников (1997) установил, что наследуемость параметров продолжительности жизни составила 80%, что характерно для признаков, детерминируемых не более чем тремя генами (Мыльников, 1997). Показано, что различия по средней продолжительности жизни и скорости старения между некоторыми линиями плодовых мух определяются одним-двумя генами, локализованными в хромосоме 2 недалеко от центромеры. Изучение вклада трех больших хромосом дрозофилы в детерминацию динамики смертности показало, что у двух пар исходных линии, различающихся по происхождению и целому ряду физиологических характеристик, динамика смертности определяется несходным образом, в то время как интенсивность перекисного окисления липидов в значительной мере обусловлена неким фактором (факторами), информация о котором содержится в третьей группе сцепления дрозофилы (Смирнова и др., 2000).
Таблица 2. Наследуемость и вариабельность продолжительности жизни у некоторых ви¬дов животных (Finch, Tanzi, 1994, с модификациями)
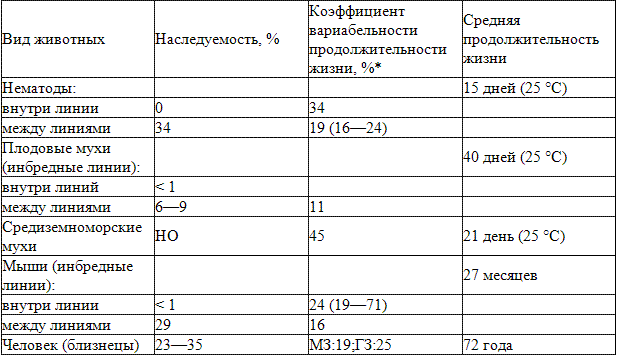
Примечание. НО - не определяли; МЗ - монозиготные; ГЗ - гетерозиготные. * Стандартное отклонение продолжительности жизни в популяции в процентах к средней продолжительности жизни.
С целью идентификации кандидатных генов в геноме лабораторных мышей несколько групп использовали так называемый анализ количества штриховых локусов (QTL-анализ). De Haan и Van Zant (1999) обнаружили на хромосоме 11 область с QTL, влияющим на продолжительность жизни, перекрывающийся с QTL, влияющим на скорость деления предшественников гематопоэтических клеток. У короткоживущих мышей линии DBA/2 (592 сут.) скорость деления этих клеток была в три раза большей, чем у долгоживущих мышей C57BL/6 (765 сут.), что авторы склонны рассматривать как in vivo эквивалент репликативного потенциала in vitro (лимита Хейфлика).
Что касается человека, то оценить наследуемость долгожительства можно, исследуя его параметры у членов одной семьи; включая приемных детей (для учета роли условий среды), а также у близнецов. Несмотря на большие трудности в интерпретации подобного материала, связанные прежде всего с существенными различиями условий жизни у представителей разных поколений и смертями от несчастных случаев, некоторые выводы все же можно сделать. В исследовании, включающем 7000 взрослых людей, дольше жили потомки родителей с большей продолжительностью жизни. Лица, чьи родители прожили более 81 года, прожили по крайней мере на 6 лет больше тех, чьи родители умерли, не дожив до 60-летия (Abbott et al., 1978).
Т. Perls и D. Terry (2203) описали 4 семьи, в которых отмечены случаи наследования исключительного долголетия. Так, в одной из семей столетние были зарегистрированы в 4 поколениях. В другой - у родителей, каждый из которых прожил более 90 лет, было 13 детей, 8 из которых прожили от 90 до 102 лет. В породнившихся двух больших семьях, имевших в двух поколениях долгожителей, в третьем поколении 23 из 46 человек достигли возраста 90-106 лет. И наконец, в потомстве родившейся в XIX веке семейной пары и имевшей 3 детей, среди 16 внуков 7 прожили более 100 лет. Расчеты ) показали, что вероятность случайного возникновения таких "кластеров" долгожительства чрезвычайно маловероятна. Все семьи жили в различных условиях, включая образ жизни и окружающую среду. Авторы полагают, что эти примеры убедительно свидетельствуют о важном роли генетических факторов в исключительном долголетии, но не обязательно свидетельствуют об одинаковой генетической его причине.
Для выявления роли вклада наследственности и факторов среды исследовали частоту различных заболеваний и смертность у приемных детей (Sorensen et al., 1986). Оказалось, что приемные дети, чьи биологические родители умерли рано от неслучайных причин, имели сами в два раза больший риск смертности от неслучайных причин. Если причиной ранней смерти одного из биологических родителей было заболевание сердца, человек, воспитанный в семье приемных родителей, все равно имел повышенный в 4 раза риск умереть от инсульта и инфаркта миокарда по сравнению с воспитывавшимися в тех же семьях биологическими детьми своих родителей. Аналогичные закономерности прослежены также в отношении смерти от инфекционных заболеваний.
Результаты большинства исследований долгожительства у близнецов свидетельствует о том, что наследуемость продолжительности жизни у человека не превышает 50% (Finch, Ruvkun, 2001). Особую значимость имеют результаты исследования шведских близнецов, которое включает большую группу близнецов, воспитывавшихся в разных семьях. Оказалось, что максимум одна треть вариабельности в общей смертности может быть обусловлена генетическими факторами, тогда как почти все остальные различия связаны с факторами окружающей среды (Ljungquist et al., 1998). Близнецы наследуют специфические гены, которые ограничивают продолжительность жизни, например определяющие предрасположенность к ожирению или атеросклерозу. Важно отметить, что различия в продолжительности жизни фибробластов монозиготных близнецов в культуре были существенно меньше таких различии между культурами фибробластов, полученных от двуяйцевых близнецов. Вместе с тем в ряде наблюдений были получены данные, свидетельствующие о существенной наследуемости долгожительства. Так, сиблинги столетних имели в 4 раза большую вероятность прожить 85 лет и более, чем сибсы тех, кто умер до 73 лет (Perls et al., 2002).
По данным Ю. П. Алтухова и В. А, Шереметевой (2000), исследовавших 15 этнических популяций в северо-евроазиатских регионах, существует значительная позитивная корреляция между гомозиготностью, определенной по 22 полиморфизмным локусам, и долголетием. К аналогичному выводу пришли М. Bonafe и соавт. (2000), обнаружившие с помощью новой inter-Alu PCR технологии увеличение гомозиготности у столетних.
См. также:
 1.2. Наследственное преждевременное старение
1.2. Наследственное преждевременное старение
1.3. Репродуктивное поведение и эволюция продолжительности жизни
1.4. Гены гибели и долголетия у круглых червей
1.5. Гены гибели и долголетия у плодовых мух
1.6. Гены долголетия у мышей
1.7. Предполагаемые гены смерти и долголетия человека
1.8. Роль специфических хромосом в старении
1.9. Изучение профиля генетического транскриптома при старении
1.10. Заключение
 Обсудить на форуме
Обсудить на форуме

