Всевластный лик, глядящий с вышины!
Настанет ночь - и взор летит из бездны,
И наши сны, взлелеянные сны
Пронизывает знанием надзвездным.
А. А. Чижевский. Космос
Наиболее существенным для живой природы явлением на Земле является смена дня и ночи, света и темноты. Вращение ее вокруг своей оси и одновременно вокруг Солнца отмеряет сутки, сезоны и годы нашей жизни. Все больше сведений накапливается и о роли эпифиза (шишковидной железы) как основного ритмоводителя функций организма. Свет угнетает продукцию и секрецию мелатонина, и поэтому его максимальный уровень в эпифизе и крови у человека и животных многих видов наблюдается в ночные часы, а минимальный - в утренние и днем. При старении функция эпифиза снижается, что проявляется прежде всего нарушением ритма секреции мелатонина, и снижением уровня его секреции (рис. 15) (Touitou, 2001; Reiter et al., 2002).
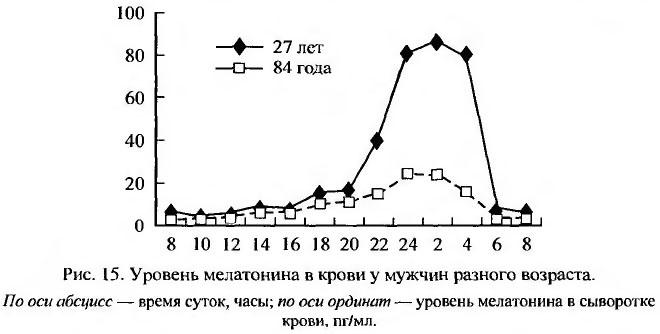
У людей в возрастной группе 60-74 года у большинства физиологических показателей наблюдается положительный фазовый сдвиг циркадного ритма (~1.5-2 часа) с его последующей десинхронизацией у лиц старше 75 лет (Gubin, Gubin, 2001). Если эпифиз уподобить биологическим часам организма, то мелатонин можно уподобить маятнику, который обеспечивает ход этих часов и снижение амплитуды которого приводит к их остановке. Пожалуй, более точно будет сравнить эпифиз с солнечными часами, в которых мелатонин играет роль тени от гномона - стержня, отбрасывающего тень от солнца. Днем солнце высоко, и тень коротка (уровень мелатонина минимален), в середине ночи - пик синтеза мелатонина эпифизом и секреции его в кровь. При этом важно то, что мелатонин имеет суточный ритм, то есть единицей его измерения является хронологический метроном - суточное вращение Земли вокруг своей оси.
Если эпифиз - солнечные часы организма, то, очевидно, любые изменения длительности светового дня должны существенным образом сказываться на его функциях и в конечном счете на скорости его старения. Циркадный ритм весьма важен не только для временной организации физиологических функций организма, но и для продолжительности его жизни. Установлено, что с возрастом нейронная активность супрахиазматического ядра снижается, при этом при содержании в условиях постоянного освещения эти нарушения развиваются быстрее (Watanabe et aL, 1995). Старые животные резистентны к действию клоргилина, стимулирующего биосинтез мелатонина в условиях круглосуточного освещения; таким же эффектом обладает разрушение супрахиазматического ядра гипоталамуса (Oxenkrug, Requintina, 1998). В ряде работ было показано, что нарушение фотопериодов может приводить к существенному уменьшению продолжительности жизни животных (Pittendrigh, Minis, 1972; Pittendrigh, Daan, 1974).
M. W. Hurd и М. R. Ralph (1998) исследовали роль циркадного ритма в старении организма на золотистых хомячках Mesocricetus auratus с мутацией ритмоводителя tau. Авторы получили 3 группы хомячков; имеющих дикий тип (+/+), гомозиготов tau-/tau- и гетерозиготов tau-/+, а затем их гибриды. Предварительные трехлетние наблюдения показали, что гетерозиготы tau-/+ имели на 20 % меньшую продолжительности жизни, чем гомозиготы. Продолжительность жизни мутантных гетерозиготов tau-/+, содержавшихся при режиме 14 часов - свет, 10 часов - темнота, была почти на 7 месяцев короче, чем в группах гомозиготов +/+ или tau-/tau- (p < 0.05), однако средняя продолжительность жизни обеих гомозиготных групп была практически одинаковой. При круглосуточном содержании хомячков в условиях постоянного слабого освещения (20- 40 люкс) с 10-недельного возраста средняя продолжительность жизни гетерозиготов и гомозиготов была одинаковой и колебалась от 15 до 18 месяцев. Для изучения причин влияния циркадного ритма на продолжительность жизни авторы имплантировали в головной мозг старых хомячков супрахиазматические ядра от плодов хомячков различного генотипа. Было установлено, что хомячки с прижившимися имплантатами жили в среднем на 4 месяца дольше, чем интактные или ложнооперированные контрольные животные. Авторы полагают, что результаты их экспериментов свидетельствуют о том, что нарушения циркадного ритма сокращают продолжительность жизни животных, тогда как их восстановление с помощью имплантации фетального супрахиазматического ядра (спонтанного осциллятора) увеличивает ее почти на 20% (рис. 16).
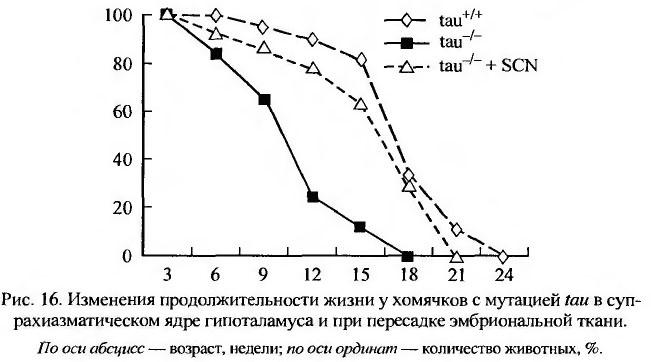
Таким же эффектом, по мнению авторов, будут обладать любые воздействия, направленные на нормализацию циркадного ритма. Интересно, что разрушение осциллятора (супрахиазматического ядра) приводит к сокращению продолжительности жизни животных (DeCoursey et al., 2000).
В 1959 г. впервые было установлено, что удаление эпифиза в молодом возрасте приводит к существенному уменьшению продолжительности жизни крыс по сравнению с контролем (Maim et a!., 1959). Эти данные были подтверждены и другими авторами (Reiter et al, 1999). В статье, опубликованной нами в 1973 г (Анисимов и др., 1973), сообщалось о том, что пептидный экстракт эпифиза (получивший позднее название эпиталамин), восстанавливает у старых самок крыс регулярные эстральные циклы и чувствительность их гипоталамических половых центров к эстрогенам - механизм, которому придается ведущая роль в возрастном выключении репродуктивной функции (Dilman, Anisimov, 1979). Тогда же нами были начаты исследования влияния этого препарата на продолжительность жизни самок крыс. Было обнаружено, что эпиталамин увеличивает длительность репродуктивного периода и на 25% - среднюю продолжительность жизни животных. Более того, он восстанавливает у части старых самок крыс способность к деторождению. Было показано также, что эпиталамин улучшает чувствительность тканей к инсулину, снижает порог чувствительности гипоталамуса к торможению глюкокортикоидами, замедляет возрастное снижение иммунных функций у животных и увеличивает продукцию вилочковой железой ее гормонов. Затем на мышах двух линий было отмечено увеличение почти на треть средней продолжительности их жизни. В этих опытах введение препарата начинали в возрасте расцвета организма, прежде всего его репродуктивной и иммунной функций - в 3-3.5-месячном возрасте, что соответствует примерно 25-30 годам человека. На мышах и крысах были проведены опыты, в которых препарат начинали вводить в возрасте, предшествующем выключению репродуктавной функции. Оказалось, что и в этом случае эпиталамин существенно увеличивал продолжительность жизни животных. Следует подчеркнуть, что во всех упомянутых наших опытах применение эпиталамина наряду с увеличением продолжительности жизни сопровождалось уменьшением частоты развития опухолей (Anisimov et al., 1994; Анисимов, 1998). Эпиталамин эффективно замедлял старение D. melanogaster (Anisimov et al., 1997).
Весьма перспективны результаты применения эпиталамина в клинической практике для профилактики преждевременного старения и развития возрастной патологии у человека (Хавинсон и соавт., 1998; Коркушко и др., 2002) (см. главу 8).
В 1987 г. Pierpaoli и Maestroni сообщили о том, что старые мыши, которым с питьевой водой на ночь давали мелатонин, жили на 20 % дольше контрольных и выглядели явно бодрее последних. Эти данные были воспроизведены на мышах различных линий и крысах, хотя геропротекторный эффект мелатонина не был одинаков и иногда отсутствовал (Анисимов, 1998; Anisimov et al., 2001). Более того, в ряде работ было установлено, что мелатонин при длительном его введении может увеличивать частоту развития новообразований (Романенко, 1983; Pierpaoli et al., 1991; Lipman et al., 1998; Anisimov et al., 2001). Подробнее данные о влиянии мелатонина и пептидных препаратов, регулирующих функцию эпифиза, на продолжительность жизни и развитие опухолей будут рассмотрены в главе 9.
Пересадка в вилочковую железу или область эпифиза старых мышей эпифиза от молодых доноров также приводила к увеличению продолжительности жизни животных (Pierpaoli, Regeson, 1994; Lesnikov, Pierpaoli, 1994).
Основные этапы изучения роли эпифиза в старении приведены в табл. 26.

Механизмы геропротскторного действия мелатонина и эпиталамина полностью не известны. Существенную роль может играть способность этих веществ угнетать свободнорадикальные процессы в организме (Reiter, 1995; Анисимов, 1998; Хавинсон и др., 199R). Как мелатонин, так и эпиталамин стимулируют клетки иммунной системы организма и замедляют старение иммунной системы, они нормализуют ряд возрастных нарушений жироуглеводного обмена, продлевают циклическую деятельность яичников у самок мышей и крыс, восстанавливают репродуктивную функцию у старых животных, обладают отчетливым антиканцерогенным эффектом (Bartsch et al., 2001; Anisimov et al., 1994, 2001c; Khavinson, 2002).
Синтез тетрапептида эпиталона (Ala-Glu-Asp-Gly) (Khavinson et al., 2000), регулирующего функцию эпифиза, открыл новые возможности в изучении роли этой железы в старении. В серии исследований изучено действие эпиталона на D. melanogaster. Пептид добавлялся в питательную среду личинок дрозофил. После этого взрослые особи, содержавшиеся без эпиталона, в течение всей жизни имели сниженный уровень продуктов перекисного окисления липидов. Удельная активность каталазы у дрозофил 14-дневного возраста была выше после воздействия на личинок эпиталоном (Khavinson, 2002). Возможно, что снижением интенсивности свободнорадикальных процессов в организме взрослых мушек, которые вывелись из получавших эпиталон личинок, отчасти объясняется увеличение их продолжительности жизни и снижение скорости старения, описанное в работах, выполненных в двух лабораториях (Anisimov et al., 1998; Khavinson et al., 2001). В то же время после воздействия мелатонином на личинок увеличение продолжительности жизни было нечетким, поскольку зависело от пола и в одной и той же линии наблюдалось в одних поколениях дрозофил, но не в других. То же относится и к ферментам антиоксидантной защиты дрозофил при действии мелатонина. Добавление мелатонина в корм взрослых дрозофил снижало показатели содержания продуктов перекисного окисления липидов и, главным образом, окислительной модификации белков. Известно, что у насекомых имеется ариламин-М-ацетилтрансфераза, необходимая для синтеза нейротрансмиттеров и меланина, который в свою очередь нужен для склеротизации кутикулы. В гомогенатах дрозофил может образовываться эндогенный мелатонин, а добавление экзогенного мелатонина в среду обитания самок дрозофил приводит к снижению копулятивной активности и отложению яиц, что теоретически должно вести к увеличению продолжительности жизни. Если предположить, что эпиталон на стадии личинок вызывает индукцию ариламин-N-ацетилтрансферазы, которая сохраняется на всю последующую жизнь, то можно объяснить и повышение устойчивости взрослых мушек к окислительному стрессу, и увеличение их продолжительности жизни, причем к этому эффекту может иметь отношение еще и склеротизация (Khavinson, 2002).
При ежедневном внутримышечном введении старым самкам макакам (20-26 лет) эпиталона в дозе 10 мкг на животное (около 0.2 мкг/кг) в течение 10 суток вечерний (21 ч) уровень мелатонина, сниженный по сравнению с имевшимся у молодых животных (6- 8 лет) в два раза, повышался в три раза, то есть оказывался выше, чем у молодых обезьян, на которых введение эпиталона в этом отношении не действовало. Кроме того, у старых обезьян наблюдалось снижение вечернего уровня кортизола и за счет этого приближение суточного профиля секреции кортизола к свойственному молодым животным (Khavinson et al., 2001). Авторы исследования предпочитают считать, что эпиталон восстанавливает возрастную чувствительность эпифиза к норадренергической стимуляции и что действие эпиталона на секрецию кортизола вторично по отношению к его эффектам на уровне эпифиза. Однако полученные ими результаты в не меньшей мере соответствуют действию эпиталона на центральные катехоламинергические функции ЦНС, коль скоро известно, что они снижаются при старении и от них зависит активация синтеза мелатонина и ингибирование секреции кортикостероидов (Анисимов, 1998). Прямых данных для выбора между двумя вышеуказанными возможностями нет. В пользу первой из возможностей свидетельствует исследование, в котором показано, что внутрибрюшинное введение эпиталона -облученным крысам сопровождается появлением ультраструктурных признаков усиления секреторной активности пинеалоцитов, нарушенной из-за облучения (Khavinson, 2002).
В пользу действия эпиталона на уровне эпифиза свидетельствует исследование на самках мышей CBА, в котором эпиталон либо дипептид Lys-Glu (вилон) вводили в дозах 0.1 мкг на животное подкожно, ежедневно, курсами по 5 дней, начиная с возраста 6 мес. и вплоть до естественной смерти животных. Параллельно еще одной группе мышей давали с питьевой водой мелатонин. Эффекты эпиталона и мелатонина по направленности совпадали для всех исследованных показателей: в обеих группах в сравнении с контролем снижались двигательная активность, длительность эстральных циклов и температура тела, не менялись поглощение корма, физическая сила, выносливость и увеличивались масса тела, интегральная оценка эффективности антиоксидантной защиты, средняя и максимальная продолжительность жизни. Единственным отличием эпиталона от мелатонина было то, что при введении эпиталона не наблюдалось повышения частоты возникновения спонтанных опухолей, которое наблюдалось при введении мелатонина (Anisimov et al., 2000, 2001). Вилон, оказывая менее выраженное геропротекторное действие, существенно отличался по паттерну от всех наблюдаемых эффектов.
Логичным представляется предположение, что мелатонин оказывает свой эффект, действуя на активность генов. В этой связи существенно, что транскрипционные факторы относятся к числу наиболее консервативных в эволюционном отношении белков, и при этом на уровне отдельных трансфакторов (даже не их комбинаций) обнаруживается тканевая специфичность в пределах организма одного вида. В частности, у млекопитающих найдена последовательность ДНК ТААТС(Т), названная "пинеальный регуляторный элемент" -- "pineal regulatory element, PIRE", повторы которой уникальны для промоторов генов, специфически экспрессирующихся в эпифизе (ариламин-N-ацетилтрасфераза, гидроксииндол-О-метилтрансфераза и эпифизарная АТФаза, ген которой мутирован при болезни Вильсона) и в сетчатке (фоторецепторы). Трансфактор из семейства гомеобоксов, специфически связывающийся с PIRE, также имеется только в эпифизе и сетчатке. Поскольку зависимость экспрессии ариламин-N-ацетилтрансферазы от цАМФ характерна только для эпифиза и сетчатки, хотя этот фермент имеется и в других тканях, предполагается, что особенности его регуляции в эпифизе обусловлены участием уникального для эпифиза и сетчатки транскрипционного регуляторного механизма.
При изучении влияния мелатонина на экспрессию генов, представленных в сердце мыши с помощью микрочиповой технологии (Анисимов С. и др., 2002) были использованы та же линия мышей (СВА). те же дозы мелатонина и схема введения, что и в опытах по изучению его влияния на показатели биологического возраста и продолжительность жизни (Anisimov et aL, 2001). В ходе эксперимента была проведена трипликативная гибридизация микрочипов с 15247 клонами кДНК с образцами ткани мышей контрольной и подопытной групп. Под воздействием мелатонина изменялась экспрессия 38 клонов. Детальный анализ функциональных подкатегорий показал, что наиболее значимыми являются изменения доли генов, имеющих отношение к регуляции клеточного цикла, и сразу двух подкатегорий генов, имеющих отношение к защитным системам клетки и организма, а именно мембранного транспорта - транспортных белков и иммунной системы, и, что особенно важно, выявлено увеличение экспрессии ряда митохондриальных генов. что согласуется с данными о его влиянии на клеточную пролиферацию, апоптоз, адгезию и его антиоксидантными свойствами (Bartsch et al, 2001; Anisimov et al, 2000; Reiter et al, 2002). Представляется весьма важным, что мелатонин стимулировал экспрессию гена лейкоза MLLT3, поскольку при длительном его введении мышам СВА наблюдалось увеличение частоты развития злокачественных лимфом (Anisimov et al., 2001). Таким образом, впервые установлено специфическое влияние мелатонина на экспрессию генов.
В целом вся совокупность описанных эффектов мелатонина более всего соответствует возможности его действия на уровне транскрипционных факторов, специфичных для эпифиза, причем это действие выражается в нормализации функций этих органов, если они нарушены в результате генетических дефектов или старения. Если это так, то перспективы профилактики и нормализации возрастных нарушений эпифиза и зависимых от него функций выглядят обнадеживающими.
См. также:
 4.1. Возрастные изменения нервной системы и старение организма
4.1. Возрастные изменения нервной системы и старение организма
4.2. Стресс и старение
4.3. Иммунологические механизмы старения
4.4. Система гормон роста-инсулинподобный фактор роста-1 (1GF-1): роль в старении и долголетии
4.5. Нейроэндокринологическая (элевационная) теория старения и формирования возрастной патологии
 Обсудить на форуме
Обсудить на форуме

