В 1978 г нами была проведена ревизия концепции Вейсманна-Свима-Хейфлика, в результате которой были сделаны следующие выводы [Гаврилов, Гаврилова, 1978, Гаврилов, Ягужинская, 1978].
1. Массовая гибель клеток в фазе II по Свиму или в фазе III по Хейфлику - это методический артефакт, обусловленный повреждением клеток в результате процедуры пересева (обработка клеток трипсином и их последующее суспендирование). Если же оценить вероятность гибели клеток вне периодов пересева, то оказывается, что вероятность смерти клеток не увеличивается с возрастом культуры, т.е. нет оснований говорить о старении на клеточном уровне.
2. Основное содержание феномена Свима-Хейфлика in vitro - это накопление в культуре постмитотических клеток, полноценных в метаболическом отношении. Процесс образования этих метаболически полноценных постмитотических клеток больше напоминает дифференцировку клеток, чем их повреждение и старение. К этому же выводу одновременно с нами пришли и другие исследователи [Bell et al, 1978].
3. Накопление в культуре постмитотических клеток не сопровождается уменьшением абсолютного числа делящихся клеток в расчете на всю образующуюся клеточную популяцию. Иначе говоря, происходит не исчезновение делящихся клеток, а лишь разбавление культуры постмитотическими клетками. Следовательно, нет никаких оснований говорить об ограниченной способности к делению всех клеток в культуре. Напротив, например, в культуре эмбриональных фибробластов человека имеются клетки, совершающие около 170 делений (вместо общепринятых 50) и обладающие свойствами стволовых клеток с неограниченной способностью к делению.
Теперь рассмотрим сделанные выводы несколько более подробно, для чего обратимся к анализу методики культивирования клеток, состоящей в следующем. Образец ткани (например, кожи) помещают в сосуд с культуральной средой и ждут, когда из этого первичного эксплантата часть клеток мигрирует на поверхность флакона и размножившись, образует сомкнутый монослой, покрывающий дно сосуда. Никто не знает при этом, сколько клеток мигрирует из трансплантата и сколько делений они совершают. По мнению Голдштейна и соавторов, эта процедура соответствует примерно 10 удвоениям популяции [Goldstein et al., 1975]. В случае же эмбриональной ткани легкого ее обрабатывают трипсином, и суспендированные клетки пересевают в сосуд с питательной средой [Hayflick, 1968]. После того как культура клеток образует монослой, ее пересевают, обрабатывая 0 1%-ным раствором трипсина, суспендируя и перенося желаемое число клеток в новые сосуды с питательной средой. Пересев в отношении 1 2 означает, что только половина клеток, образовавших сомкнутый монослой, пересевается в сосуд с той же поверхностью. Когда и в этом сосуде в результате деления клеток образуется монослой, считают, что произошло удвоение клеточной популяции, соответствующее в среднем одному делению клеток. Подобную операцию повторяют многократно (в случае эмбриональных фибробластов человека около 50 раз), до тех пор, пока культура не потеряет способности к быстрому восстановлению своей численности. Теперь попытаемся оценить число делений, которое совершают клетки. Известно, что через сутки после операции пересева в живых остается только 50-70% клеток "молодых" культур и лишь 25% клеток "старых" культур (эти клетки являются более крупными и сильнее всего повреждаются при суспендировании) [Гаврилов, Гаврилова, 1978, 1982].
Примечание: Такое массовое разрушение клеток при пересеве в сочетании с низкими темпами восстановления численности "старых" культур и порождало иллюзию естественной массовой дегенерации и гибели клеток описанную в ряде работ [Swim, 1959, Hayflick. Moorhead, 1961, Hayfhck, 1968]
Таким образом. 40-60 пересевов соответствуют не 40-60 удвоениям популяции, а 80-120 удвоениям, которые совершают выжившие клетки. Далее, одно удвоение клеточной популяции соответствует одному делению клеток лишь в том случае, если все клетки способны делиться. На самом же деле доля делящихся клеток сильно уменьшается при культивировании и к последнему пересеву составляет всего 10-20% [Там же]. Можно показать, что в том случае, когда только 10% клеток сохраняет способность к делению, им необходимо поделиться 10 раз, чтобы численность всей культуры удвоилась [Гаврилов, Ягужинский, 1978, Гаврилов, Гаврилова, 1982]. Если учесть это обстоятельство, то окажется, что 80-120 удвоений численности клеток соответствуют 170±30 делениям. Итак, оказывается, что 50 пересевов клеток человека соответствует не 50, а 170±30 делениям.
Самое главное, однако, состоит в том, что и это число делений оказывается не окончательным. Дело в том, что гибель клеточной культуры определяется весьма своеобразно. Оказывается, культуру клеток называют мертвой лишь потому, что в течение произвольно заданного интервала времени (обычно от одной до четырех недель) ее численность не возрастает до желаемой величины (обычно задается численность, в два-четыре раза превышающая исходную).
Примечание: В качестве примера приведем следующее определение: "Культуры считались мертвыми, когда они не могли достигнуть сомкнутого монослоя после 4 недель при еженедельной смене среды" [Goldstein, Singal, 1974, р. 360).
Более того, в "мертвых" культурах еще остается 10-20% клеток, способных делиться, поэтому культура может компенсировать естественную гибель клеток (которая, кстати, не увеличивается) и даже медленно расти [Гаврилов, Гаврилова, 1978; 1982]. Оказалось также, что абсолютное число делящихся клеток в расчете на всю образующуюся клеточную массу не уменьшается, а происходит лишь их разбавление неделящимися клетками. В результате, когда доля делящихся клеток становится близка к 10%, культура действительно не может расти столь быстро, чтобы ее считали живой. Таким образом, феномен "гибели" культуры клеток не только не эквивалентен гибели клеток, но даже не означает того, что все клетки культуры имеют ограниченную способность к делению. В результате проведенных исследований нами был сделан вывод, что в культуре клеток происходит их дифференцировка с образованием неделящихся клеток, но некоторые клетки культуры являются стволовыми и могут размножаться неограниченно [Гаврилов, Ягужинский, 1978].
Некоторые из сделанных нами выводов были одновременно и независимо подтверждены другими авторами [Belletal., 1978, р. 1160]:
"...одной из наименее документированных особенностей гипотезы старения in vitro является общепринятое утверждение о том, что клетки в фазе 3 являются нежизнеспособными и в конце концов погибают. Это утверждение не имеет никакого серьезного экспериментального подтверждения ...Наш опыт свидетельствует о том, что сам по себе пересев неизбежно приводит к потере клеток. ...Мы наблюдали выживаемость фибробластов крайней плоти (штамм 1519) в культуре в течение длительного периода после того, как клеточное деление прекращалось. Наши первые культуры клеток штамма 1519 поддерживались живыми в среде Хэма ...в течение 22 месяцев и в течение периода от года до 14 месяцев после прекращения клеточных делений ...Биохимические исследования показали, что, несмотря на то, что клетки фазы 3 отличаются в некоторых отношениях от клеток фазы 2, они, по существу, нормальны. ...Сообщения о биохимических различиях между клетками в фазе 2 и фазе 3 отражают различия в дифференцировке, а не в возрасте".
Примечание: Фазы 2 и 3 в данной статье соответствуют классификации Хейфлика, а не Сейма. Повидимому, авторы данной статьи, так же как и многие другие исследователи, просто не знакомы с основополагающими работами Сейма и поэтому его статей не цитируют.
Впоследствии и сам Л. Хейфлик совместно с Т. Матсумурой и 3. Зеррудо [Matsumura et al., 1979] подтвердил отсутствие массовой гибели клеток в фазе III по его классификации. Авторы этой работы в течение шести месяцев поддерживали "мертвые" культуры, некоторые из которых за это время в 16 раз увеличили свою численность. Никаких признаков приближающейся массовой гибели клеток обнаружить не удалось, хотя срок наблюдения за "мертвыми" культурами иногда превышал даже год (для сравнения отметим, что вся предшествующая процедура 50 пересевов занимает обычно шесть-восемь месяцев) [Hayflick, 1968].
В результате этих исследований Хейфлик изменил свое представление об ограниченной продолжительности жизни клеточных культур. Первоначально он излагал свои результаты следующим образом (рис. 54):
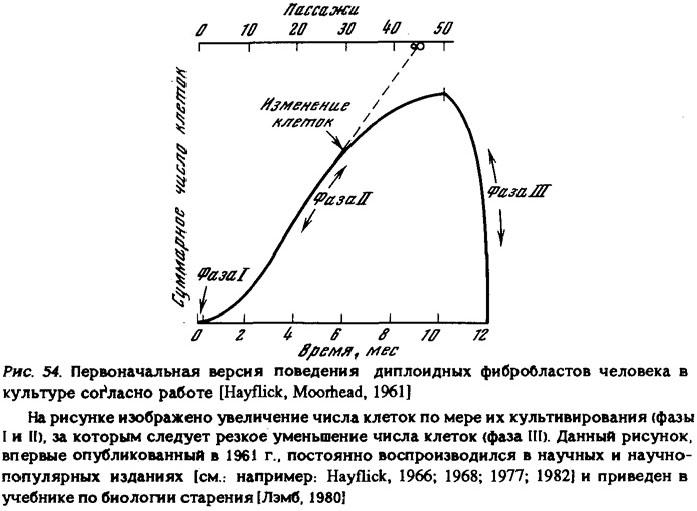
"Мы обнаружили, что фибробласты, взятые от четырехмесячного человеческого эмбриона, удваивались таким образом в числе в среднем 50 раз (от 40 до 60). Истощив свою способность к делению, клетки гибли" [Hayflick, 1968, р. 35]. В работе 1979 г. приведена уже принципиально иная схема (рис. 55)
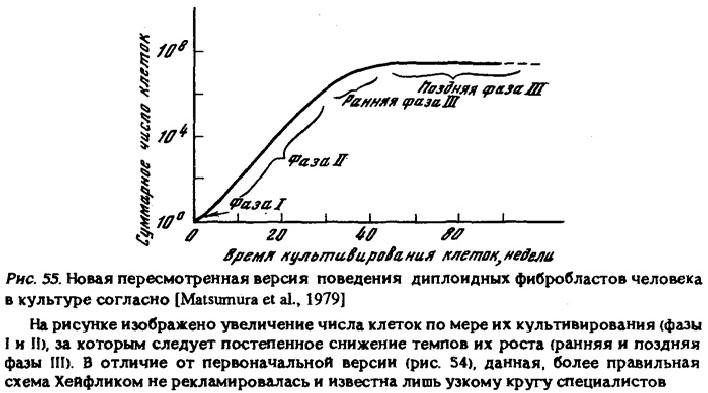
и все излагается иначе: "В ранней фазе III культура еще пролиферирует, хотя скорость пролиферации снижается. В последней фазе III клеточная пролиферация очень низка... В течение этого периода не наблюдалось ни признаков окончательной смерти культуры, ни спонтанного приобретения неограниченного пролиферативного потенциала" [Matsumura et al., 1979, р. 332, ЗЗЗ]. Следует подчеркнуть, что изменение позиции исследователя делает ему честь как ученому, поскольку цель науки состоит не в отстаивании догматов, а в поиске истины. Однако достойно сожаления то, что вскрывшаяся истина стала известна лишь очень узкому кругу специалистов, а в таких ведущих научно-популярных журналах, как "Scientific American", продолжалась реклама старой легенды, что "в конце концов клетки претерпевают различные дегенеративные изменения и гибнут" [Hayflick, 1980, р. 42]. Конечно, удивительно, что пропаганда этого мифа велась тем же автором, который годом ранее так хорошо показал его несостоятельность, однако эта загадка выходит за пределы тематики данной книги.
Примечание: Еще более удивительно, что тот же автор три года спустя опять публикует устаревшую схему (рис. 54) вместо им же исправленной схемы (рис. 55) и вновь излагает опровергнутую версию, что "после исчезновения митотической активности культура полностью дегенерирует" [Hayflick, 1982, р 641].
В то же время становится понятен механизм канонизации устаревших представлений о пределе клеточных делений в современных томах таких солидных изданий, как, например, "Британская энциклопедия".
См. также:
 Лимит Хейфлика, открытие
Лимит Хейфлика, открытие
 Ген смерти
Ген смерти
Укорочение теломеры. Возрастное исчерпание генетического кода
 Обсудить на форуме
Обсудить на форуме

