Линия SAM (senescence accelerated mouse) была получена путем селекции мышей линии AKR/J (Takeda et a)., 1997; Takeda, 1999; Hosokawa, 2002). Существует несколько сублиний мышей, живущих в среднем 12-15 месяцев, предрасположенных к ускоренному старению, - SAMP (prone), и устойчивых к преждевременному старению - SAMR (resistant), являющихся контролем к SAMP. Все эти сублинии довольно близки генетически, но отличны от линии AKR/J (Takeda, 1999). Данные генотипирования с использованием микросателлитных маркеров позволили предположить, что четыре локуса, расположенных в хромосомах 4, 14, 16 и 17, содержат гены, ответственные за ускоренное старение у этих мышей. Показано, что мыши SAMP нормально развиваются до 4 месяцев, затем имеют признаки ускоренного старения: потерю волос, кожные изъязвления, уменьшение локомоторной активности, ухудшение памяти и способности к обучению, эмоциональную дезориентацию, нарушенным циркадный ритм, атрофию легких, повреждения сердца, катаракту, увеличение продукции активных форм кислорода (АФК) и уровня 8-оксигуанина во всех органах (Takeda et al., 1997; Takeda, 1999; Юнева и др., 2000). Количество Си,Zn-супероксиддисмутазы (СОД) в митохондриях у мышей SAMP1 в 2 раза меньше, чем у мышей линии SAMR1 (Park et al., 1996). Имеются данные о повышенном уровне перекисного окисления липидов в различных органах мышей SAMP по сравнению с таковым у линий SAMR. Hosokawa (2002) полагает, что у мышей SAMP имеют место нарушения в системе переноса электронов в митохондриях. Продолжительность репродуктивного периода у мышей линии SAMP короче, чем у SAMR, старение репродуктивной функции ускорено по сравнению с таковой у SAMR (Miyamoto et al., 1995).
В табл. 40 приведены сведения об основных нарушениях функций у мышей SAM. Hosokawa (2002) склонен рассматривать мышей SAM скорее как модель "ускоренного старения", а не как модель "преждевременного старения".

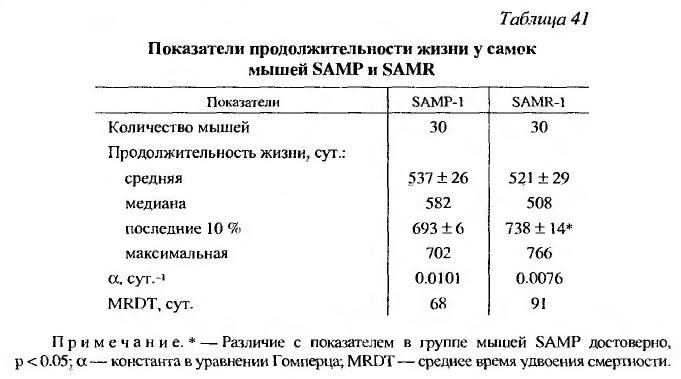
По данным Е. Е. Егорова и соавт. (Yegorov et al., 2001), максимальная интенсивность смертности мышей SAMP-1 наблюдалась в возрасте 10-12 месяцев, тогда как у SAMR-1 - в 14--16 месяцев. Максимальная продолжительность жизни у мышей этих сублиний составила 16 и 23 месяца, В наших наблюдениях мыши SAMP-1 и SAMR-1, полученные нами, как и в опытах Е. Е. Егорова из Московского государственного университета, и размножавшиеся в условиях нашего вивария жили дольше, чем в их работе, причем, не наблюдалось различий в средней продолжительности их жизни. Однако 10% максимально проживших животных, максимальная продолжительность жизни и среднее время удвоения смертности были меньше, а скорость старения популяции (а) - больше у мышей SAMP-1 по сравнению с их диким типом SAMR-1 (табл. 41).
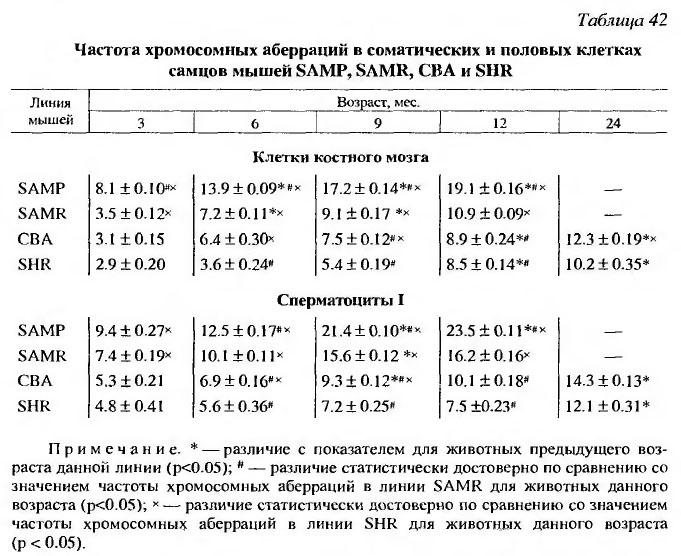
Активность О6-метилгуанин-ДНК метилтрансферазы - фермента, репарирущего ДНК при алкилировании гуанина в О6 положении, у мышей SAMP1 такая же, как и у SAMR1. У мышей с ускоренным старением линии SAMP1 наблюдали увеличение частоты хромосомных аберраций в возрасте от 3 до 8 месяцев, в то время как у линии SAMR1 выявляется только незначительное ее увеличение (Hosokawa, 2002). Отмечено увеличение с возрастом накопления в клетках мышей линии SAMP микроядрышек по сравнению с линией SAMR (Урываева и др., 1999). Возрастные изменения частоты соматических Hprt мутаций в лимфоцитах селезенки и накопление повреждений в ДНК (главным образом, одноцепочечных разрывов) развивались быстрее в шести изученных органах у мышей SAMP1, по сравнению с линией SAMR1 (Odagiri et al., 1998; Hosokawa et al., 2000). Частота хромосомных аберраций в клетках костного мозга у мышей SAMP-1 нарастала быстрее, чем у мышей линии SAMR-1, СВА и SHR (Ршенфельд и др., 2002) (табл. 42).
Первичные культуры фибробластов, полученные от мышей SAMP-1, в среднем проходили in vitro меньшее число пассажей (8.7 удвоений), чем полученные от мышей SAMR-1 (12.3 удвоений), тогда как фибробласты мышей линии СВА проходили в среднем 20 удвоений. Непролиферирующие старые фибробласты SAM жили в культуре 100-150 дней, тогда как старые фибробласты мышей линии СВА - 220-260 дней (Yegorov et al., 2001). Уменьшение выживаемости фибробластов мышей SAM сопровождалось ускоренным накоплением в них клеток, в которых выявлялась -галактозидаза. Длина теломер у мышей SAMP-1 и SAMR-1 была одинаковой, но у ускоренно стареющих мышей SAMP-1 наблюдалась большая гетерогенность длины теломер. Активность теломеразы была выше в эмбриональных фибробластах мышей SAM по сравнению с таковой в фибробластах долгоживущих мышей линии СВА (Yegorov et al., 2001).
Частота развития спонтанных лимфом составляла 17.5 % у различный сублиний мышей SAMP (от 0 % у SAMP11 и SAMP6 до 60.2% у SAMP7) и 13.7% у различный сублиний мышей SAMR (2.7 % у SAMR5 и 23.1 % у SAMR4). Частота злокачественных опухолей других локализаций составляла от 0 до 4.8 % у мышей SAMP и от 3.8 до 4.1 % у SAMR (Takeda et al., 1997). По нашим данным, у мышей SAMP-1 и SAMR-1 частота спонтанных лимфом не различалась и составляла 67-71% (Розенфельд и др., 2002). Титр вируса лейкемии был б льшим в крови и селезенке и намного больше в мозге мышей SAMP8, чем в таких же тканях у мышей линии SAMP1 (Meeker, Carp, 1997). Sugimura et al. (1994) обнаружил у мышей SAMP высокую частоту стромальной гиперплазии с фиброзом и воспалением в задней доле предстательной железы. Атипичные железистые эпителиальные клетки и криброзную деформацию желез наблюдались в задней боковой доле предстательной железы мышей линии SAMP.
См. также:
 6.4.1. Трансгенные мыши с суперэкспрессией гормона роста
6.4.1. Трансгенные мыши с суперэкспрессией гормона роста
6.4.2. Мыши с генетическим ожирением
6.4.4. Мыши с мутацией гена klotho
6.4.5. Трансгенные модели для изучения функции генов репарации ДНК
6.4.6. Трансгенные мыши с суперэкспрессией гена Си, Zn-супероксид дисмутазы
6.4.7. Мутационные и трансгенные модели иммуностарения
6.4.8. Трансгенные модели возрастных нейродегенеративных заболеваний
6.4.9. Мыши с нокаутным геном белка р53
6.4.10. Регуляция межклеточного взаимодействия и трансгенные модели
6.4.11. Теломераза: трансгенные и нокаутные мыши
...
 Обсудить на форуме
Обсудить на форуме

