Бессмертная линия стволовых клеток остается наилучшим лабораторным GМP- сырьем для биотрансплантатов в отсутствии фетальной ткани. Стандартно получаемые клетки из нестандартного биосырья незаменимы на стадии предклинических испытаний. Международные GМP-стандарты отдают предпочтение генетически однородным клеткам (Snyder E.,1994 ). (Таб 1) . Уже много десятилетий линии пассируемых плюрипотентных раковых клеток использовались для трансплантаций на животных. Наибольшее распространение получила линия РС12 , выделенная из феохромацитомы человека. Эти пассируемые мультипотентные предшественники являются клетками-дериватами нервного гребня. Линия CG9 была получена из глиомы человека . Геном клеток этой линии наделен плюрипотентностью, как и геном тератокарциномы Р19 . Но клетки этих линий не встриваются в бластоцисту и не интегрируются в нормальное развитие зародышей в отличие от других линий Эск.
Таблица 1 Преимущества GLP- линий Нск для предклинических испытаний:
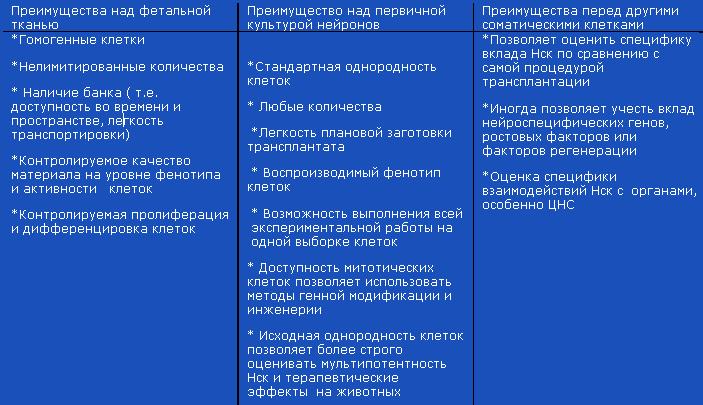
Линии Нск особенно важны в предклинических исследованиях на животных, имея превосходство перед гетерогенной первичной культурой . Без линий было бы невозможно выявить все возможные пути воздействия клеточного трансплантата на поврежденный мозг. Большой интерес вызвала условно бессмертная линия Нстволовые клетки, изолированная из нейроэпителия трансгенных зародышей мыши. Для иммортализации использовали температуро-чувствительный онкоген. В культуре при 20-25С клетки неограниченно пролиферировали, но переставили делиться и дифференцировались при 37С (Hodges H., Veizovoc, Bray R. еt al., 2000). Главное преимущество линейных клеток для трансплантации обусловлено их тремя свойствами: 1) высокая миграционная активность 2) дифференцировка в нейроны с разным профилем медиатора, причем окончательная дифференцировка донорских клеток дИнститут Высоких МЕДИЦИНСКИХ Технологийовалась микроокружением. Следовательно, общий пул клеток, трансплантированных в латеральный желудочек, превращался в разные нейроны в разных отделах мозга реципиента 3) в зоне окончательной локализации пересаженные клетки не нарушали нормальной организации ткани, не вызывали локальной дисплазии или гибели клеток. Мышиная линия иммортализированных прогениторных клеток МНР-36, полученная на фирме ReNeuron, испытывалась на крысах с постишемическим поражением мозга, вызванными краткосрочным пережатием средней церебральной артерии (Veizovic T., Beech J.S., Stroerner R.P. et al., 2001). Нарушение двигательных навыков оценивали рядом объективных количественных тестов, в том числе по стволовые клеткиорости/эффективности снятия с лап кусочков липкого пластыря. Если МНР-36 клетки трансплантировали между 1-2-й нед после ишемии, удавалось компенсировать ранние нарушения двигательных расстройств, которые никак не могли объясняться встраиванием клеточного трансплантата в мозговую ткань крысы ( слишком короткие сроки для клеточного замещения утраченных нейронов). Одностроннее введение однородных прогениторных клеток давало коррецию повреждения в обоих полушариях, что говорило о стимуляции регенерации или формировании новых зон синаптогенеза, с помощью которых компенсировались утраченные двигательные навыки. Стереотаксические пересадки линии МНР-36 в гиипокамп старых крыс или взрослых крыс с повреждением СА1 зоны гиппокампа ( модель утери пространственных навыков и переобучения) были использованы для доказательства возможной коменсации утраченных функций гиппокампа за счет образования нового химерного органа, собранного из нейронов донора/реципиента (Hodges R., Veizovic T.,Bray N. еt al., 2000). Модель использована также для региональных трансплантаций Нск с целью компенсации феноменов “старения” мозга.
В другой работе ( Philips M.F., Mattiasson G.,Weloch T. et al, 2001) прогениторные клетки линии HiB5 , изолированные из гиипокампа и трансфицированные геном ростового фактора NGF, вводили через 48 часов после механической травмы головного мозга. Уже через 3-5 суток ( т.е. в очень ранний посттравматический период) отмечали значительную компенсацию нарушенных моторных функций животного. Быстро развивающийся положительный эффект невозможно объяснить на основании известных клеточных механизмов компенсации, поскольку все они требовали значительного времени.
В совместном шведско-американском проекте, вполненном в лаборатории Рона МакКея и Эндерса Бьорклунда, были получены температуро-зависимые линии Нск/прогениторных клеток, изолированные из гиппокампа и стриатума мозга человека ( Lundberg C., Martinez-Serrano A., Cattaneo E. еt al., 1997). Если клетки гиппокампа активно обновлялись в мозге взрослых животных и человека, то стриатум практически не имел детектируемого обновления клеток. Поэтому было важно сопоставить эффекты пересадок прогениторных клеток в гиппокамп и стриатум. В стриатуме взрослых животных не бывает обновления собственных клеток. Поэтому БУДР селективно визуализировал донорские клетки in situ. После пересадки клетки выживали в течение 6 мес. Количество живых клеток в трансплантате было достоверно больше (в 2-3 раза) при пересадке в мозг животного с экспериментальным повреждением стриатум. Однако обнаружить встраивания донорских клеток в сети нейронов ткани реципиента не удалось. Поэтому пересадки лабораторных линий прогениторных клеток предлагалось использовать для направленной доставки генов, контролирующих выработку нейротрофических факторов или митогенов, стимулирующих регенерацию собственной паренхимы.
Многократно пассируемую линию Нск получил в Италии Анджело Вескови, не прибегая к онкогенам. (Kallos M.S., Behie L.A., Vescovi A.L.,1999). Коммерческие линии Нск человека в суспензионных биореакторах были получены в совместном итало-канадском проекте ( Kallos M.S., Behie L.A.,1999). Собственные линии Нск человека для экспериментов на животных, а также для подготовки первых клинических испытаний получили испанские исследователи ( Rubio F.J., Bueno C.,Villa A. et al, 2000; Villa A., Snyder E.Y.,Veskovi A.L. et al., 2000 ). В этих работах использовали селекционное пассирование клонов Нск в среде, содержащей сразу два ростовых фактора (EGF и bFGF). Среди первичных нейросфер удалось наткнуться на клоны с необычно мощной и долгосрочной потенцией к самообновлению и высоким индексом пролиферации. Полученную линию Нск пассировали более 50 раз в течение 1,5 лет. От нее получено уже сотни млн прогениторных клеток для биотрансплнататов.
Линию плюрипотентных Нск мозжечка зародышей мыши (С17-2) получил Evan Snyder для лечения модельных заболеваний мышей, в том числе экспериментальной демиелинизации ( Taylor R.M.,Snyder E.Y.,1997). У 60 % мышей линии shiverer трансплантация Нск вела к устранению тремора - главного признака демиелинизации. При введении в мозг плюрипотентные клетки этой линии мигрировали и расселялись практически по всем регионам мозга. (Yandava B.,Billinghurst L.,Snyder E.Y., 1999). Существенно, что стволовые клетки наделены автоматикой цленаправленной миграции в зоны повреждения мозговой ткани. Детали этих исследований можно найти в патенте Снайдера ( No 5958767). Пересадки линии Нск человека, как и мышиной линии С17-2 использовали для доказательства терапевтической эффективности клеток с целью лечения наследственных заболеваний ЦНС. Например, пересадки Нск в мозг мышей с накаутом гексаминидазы ( модель болезни Тей-Сакса у детей) приводили к существенному снижению уровня токсичных Gm2 -ганглиозидов в реципиентной ткани мозга за счет доставки лимитирующего фермента (гексаминидазы) лизосом, удаляющего избыток накопленных фосфолипидов ( Lacorazza H.D., Flax J.D., Snyder E.Y. et al.,1996). Ранее было известно, что пересадки донорского костного мозга были эффективны в ликвидации последствий болезни в висцеральных органах, но не в ЦНС. Пересадки линий Нск были предложены в качестве прямой доставки нормальной аллели гена гексаминидазы для корррекции метаболизма фосфолипидов в головном мозге. Платформу будущей терапии формирует доказанная клеточная стабильная химеризация мозга реципиента донорскими Нск.
Линии Нск были изолированы не только из зародышей, но новорожденных и взрослых мышей ( Flax J.D.,Aurora S.,Yang C. еt al, 1998). По той же стратегии была выделена Нск из перивентрикулярной ткани фетального мозга. Сперва получали первичную клоногенную культуру Нстволовые клетки, затем отбирали наиболее быстро пролиферирующие клоны, которые трансфицировали конструкцией из рестровирусного вектора и двух генов : v-myc ( для иммортализации) и lac-z ( микробный ген бета-галактозидазы для визуализации ). Первый ген был поставлен под промотер нестина, второй - под промотер неомицина. З линии были получены из трех разных быстро растущих клонов. Клональность линии подтверждалась единственным местом вставки v-myc в геном. Линии длительно пассировали без изменения генотипа (анеуплоидия) и фенотипа (маркеры Нск). Сохранялась плюрипотентность клеток in vitro. У линий не была выявлена туморигенность. Важнейшей характеристикой наиболее изученных линий Нск оставалась пластичность: большинство трансплантируемых клеток дифференцировались в нейроны и глию, преобладающие в месте нахождения трансплантата. Вновь подтвердилась первостепенная значимость сигналов микроокружения в судьбе клеточного трансплантата (Vescovi A.L., Snyder E.Y., 1999). Первые заметные результаты эта лаборатория получила, используя пересадку линии 17.2 в мозжечок мутантным мышам meander tail. Эти животные имели наследственные дефекты костной системы и аномалии архитектоники в мозжечке. Основу мозжечковых расстройств составляли дефекты миграции незрелых гранулярных клеток. Это вело к двигательным расстройствам. Пересадки клеток линии 17.2 восстанавливали клеточную организацию внутреннего и внешнего гранулярного слоя. Пересадки позволили выяснить как функционирует мутантный mea ген в мозжечке, путем сопоставления коррекции мозжечковых расстройств с гистологической картиной поэтапного восстановления цитослоев в мозжечке ( Rosario C.M., Yandava B.D., Kosaras B. et al., 1997). При трансплантации в боковые желудочки или субвентрикулярное пространство мозга новорожденных клетки активно мигрировали в разные отделы мозга по преформированным путям. Трансплантация этих клеток в герментативный слой мозжечка приводила к миграции, дифференцировке и встраиванию прогениторных клеток в наружний гранулярный слой. Реже из донорских клеток формировались клетки Пуркинье и олигодендроциты. Только некоторые линии Нск отличались высокой стволовые клеткиоростью миграции и высоким индексом встраивания в преформированный гранулярный слой. Пересадки первичной суспензии клеток фетального мозжечка в мозжечок иммунодефицитных животных были более эффективными в терминах количества и эффективности восстановления цитоархитектоники слоев донорскими клетками (Pundt L.L., Jorn E.A., Low W.C., 1997). Возможно, что причиной высокой приживляемости клеток является исходный синергизм взаимодействия гетерогенных прогениторных/бластных клеток, созревающих в разном направлении. Функциональное встраивание донорских клеток доказывалось необратимой утерей экспрессии (v-myc) и клеточных делений. Репаративные способности этой линии были сперва апробированы на линии мутантных мышей meander, имевших недоразвитие гранулярного слоя нейронов в передней доле мозжечка. Введение донорских Нск в паренхиму вело к направленной миграции и аккумуляции прогениторных клеток вдоль границы внутреннего гранулярного слоя, где завершалась их дифференцировка. Донорские клетки после встраивания визуализировали с помощью маркерной бета-галактозидазы. Недостатком линий в плане клинического применения является малый процент новообразованных нейронов по отношению к глии. Это происходило в том числе из-за высокой концентрации ВМР-2/4 в паренхиме взрослого мозга ( Gokhan S., Song Q., Mehler M.F., 1998).
Весьма перспективными представляются пересадки Нск для лечения атаксии-телангиоэктазии - фатального наследственного заболевания детей. Болезнь вызывается мутацией АТМ (АТ-Mutated) гена, контролирующего синтез сигнальной фосфоинозитолкиназы (PI3-kinase), участвующей в функционировании мультиферментного сигнального комплекса в хроматине. Этот сигнальный комплекс имеет множественные функции в клетке. Плейотропные эффекты мутации АТМ проявляются в гибели клеток Пуркинье мозжечка, дегенерации мотонейронов, иммунодефиците из-за дефектов стромы тимуса ( максимальное содержание АТМ комплекса обнаружено в мезенхимальных клетках). На клеточном уровне дефект проявляется в ускоренной гибели клеток вследствие утери чувствительности к кислородным радикалам. Утрата иммунитета ведет к раннему возникновению опухолей. Пересадки Нск в мозжечок РСD(Purkinje cell deficient) мышей, приводили к восстановлению слоя клеток Пуркинье , формированию функционального слоя донорских клеток, контактирующих с клетками внутренннего гранулярного слоя реципиента. Морфологические признаки восстановления клеточных слоев сопровождались уменьшением тремора и патологии движений.
Похожие на атаксию мозжечковые расстройства вызываются генетическим дефицитом активатора плазминогена. В мозговой ткани зародыша активатор плазминогена контролирует направленную миграцию нейронов и рост нейритов. Его дефицит особенно проявляется дезорганизацией роста клеточных слоев в мозжечке с симптомами атаксии (Seeds N.W.,Basham M.E.,Haffke S.P.,1999).
До 40% пациентов с острым инфарктом миокарда и нарушением системного кровообращения имели нарушения памяти и распознавания новизны в результате ишемического повреждения гиппокампа (Virley D.,Ridley R.M., Sinden J.D. et al.,1999). Близкие нарушения памяти и обучения имели пациенты с болезнью Алцхеймера на базе нейродегенеративных нарушений гиппокампа. Глобальная ишемия в посоперационном периоде вследствие гипоперфузии мозга также является частой причиной ишемических повреждений гиппокампа и других мозговых структур. Могут ли такие поздние локальные повреждения нейронов взрослого мозга «опознаваться» донорскими Нск? Для этого разработали модель избирательного повреждения пирамидальных нейронов двигательной коры взрослых мышей с помощью направленного фотолиза. Сперва к клеткам ограниченной зоны доставляли фотосенсибилизатор на гранулах микроносителя. На втором этапе зону мозга освещали лазерным лучем, что через 8-10 дней вызывало массовую локальную гибель клеток апоптозом (Snyder E.Y., Yoon C., Flax J.D. et al.,1997). Затем Нск линии 17.2, меченые геном lacZ, вводили в латеральный желудочек . Было показано, что основная часть пересаженных клеток мигрировала и накапливалась в зоне повреждения. Характерно, что интенсивная направленная миграция незрелых клеток происходила без формирования радиальной глии, т.е. эта миграция клеток не повторяла события эмбриогенеза во взрослом мозге животного. При пересадке нейросфер миграция осуществлялась с помощью клеток радиальной глии, которые опережающе возникали из клонов Нск.
Интерес в последние годы вызывают некоторые Нох-гены в связи с проблемами иммортализации стволовых/прогениторных клеток ЦНС. Была обнаружена асимметрично высокая экспрессия гена Рах-3 в медуллосаркоме человека. В изолированной линии медуллосаркомы с высокой экспрессии Рах-3 были отмечены следующие общие изменения фенотипа клеток: а) 2-4-кратное повышение активности 8-полисиалилтрансферазы, генерирующей pоly-S-NCAM на поверхности клеток б) клетки увеличивают миграцию за счет утери межклеточных контактов ( Mayanil C.S., George D.,Mania-Farnell D., 2000). Далее у многих миогенных и нейрогенных опухолей была выявлена повышенная экспрессия генов Рах-з/Рах-7. Эта фенотипическая особенность сохранялась в клеточных линиях, изолированных из первичной опухоли ( Barr F.G.,Fitzgerald J.C., Ginsberg J.P.,1999).
Нск можно идентифицировать по поведению в реципиентной ткани мозга зародышей и постнатального мозга. Согласно многочисленным данным, Нск и прогениторные популяции при введении в полость желудочков активно мигрировали в паренхиму, накапливались в субвентрикулярной области. Далее клетки расселялись буквально по всем отделам развивающегося мозга - от коры больших полушарий и мозжечка до ствола мозга (Brustle O.,Choudry K., Karram K. et al., 1998). Идентификацию Нск человека, расселяющихся в мозге развивающихся зародышей крыс, проводили до и после рождения животных с помощью генетических маркеров ДНК человека (сиквенсы alu) и иммуноморфологических маркеров. Пересадки мышиных Нск в развивающийся мозг зародышей крыс позволили без иммуносупрессии получать новорожденных животных, мозг которых собран из клеток двух видов грызунов (Olsson M., Bjerregaard K., Winkler C. et al., 1998). Донорские ростки стволовых клеток не только делились и мигрировали с высокой стволовые клеткиоростью, но и обладали способностью находить места повреждения паренхимы мозга. Например, пересадки нормальных Нск в мозг животных с наследственными аномалиями приводили к частичной компенсации функционального дефекта. Терапевтический эффект обусловлен стабильным выживанием и функционированием донорских нейронов, возникших из стволовых клеток в нервной ткани реципиента (Yandave B.D., Billinghurst L.L., Snyder E.,1999; Zigova T., Sanberg P.R., 1998). Трансплантация предшественников олигодендроцитов в латеральный желудочек приводила к массированной миграции донорских клеток по всей паренхиме мозга животных-реципиентов с появлением множественных очагов ремиелинизации (Learish R.D., Brustle O., Zhang S.C. et al., 1999).
Пересадки Нск на животных подтвердили самое важное : трансплантированные в паренхиму мозга незрелые клетки вели себя предсказуемо, не формировали аномальных клеток, не генерировали опухолей (Zhang S.C., Wernig M., Thomson J.A.,2001) После пересадки Нск/прогениторные популяции продуцировали только нейроны, олигодендроциты, астроглию, но не гематогенные ростки, кардиомиоциты, или клетки серкреторных желез. Факторы и сигналы окружающей среды in vivo оказывали предпочтительное влияние на дифференцировку имплантированных незрелых клеток. Это позволяет рассчитывать на безопасность метода в клинике .
Сравнительный анализ выживаемости, пролиферации, стабильной дифференцировки Нск in situ, полученных из разных источников, четко доказал преимущество: а) прогениторных клеток поликлонального происхождения ( более полиморфных популяций) б) клонов (нейросфер) по сравнению с культурой монодисперсных клеток, растущих прикрепленным монослоем ( см. детали в статье Rubio F.J., Bueno C/. Villa A. et al.,2000).
См. также:
 6. Регионализация и сегментация нервной трубки
6. Регионализация и сегментация нервной трубки
7. Первичный нейро - и глиогенез
8. Направленная миграция прогениторных клеток: взаимодействие с радиальной глией
9. Нейрональные стволовые клетки in vitro
10. Методические трудности получения клонов Нск из Эск
11. Получение нейронов из Эск
13. Трансплантация Нск/прогениторных клеток в развивающийся мозг эмбрионов
14. Трансдифференцировка Нск после трансплантации
15. Нейромезенхимальные стволовые клетки нервного гребня
16. Литература
...
 Обсудить на форуме
Обсудить на форуме

