Представление о том, что старение сопровождается прогрессивным снижением интенсивности секреции всех гормонов, падением их концентрации в крови, неверно. Оказалось, что содержание одних гормонов к старости падает (к примеру, половые стероиды, тиреоидные гормоны), других существенно не изменяется (альдостерон, кортикостерон), третьих выраженно растет (ФСГ, ЛГ, вазопрессин). Многие противоречия по поводу возрастных изменений концентрации гормонов связаны с использованием различных методов их определения, с сопоставлением несовпадающих возрастных групп, с видовыми особенностями возрастных изменений секреции и др. Для старения секреторных элементов характерна гетерохронность и гетерогопносгь. Так, возрастные изменения в пучковой зоне коры надпочечников выражены больше, чем в сетчатой; в одной и той же железе (кора надпочечников, щитовидная железа и др.) наряду с атрофированными, структурноповрежденными секреторными элементами находятся клетки в состоянии гипертрофии и гиперфункции. Это определенный адаптивный механизм, направленный на поддержание исходного уровня секреции.
Таблица 2. Содержание ТТГ в аденогипофизе, а также ТТГ, тироксина (T4), трийодтиронина (T3) в крови у крыс разного возраста
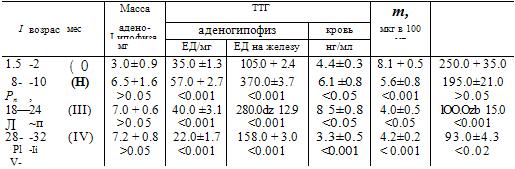
Примечание Римские цифры - возрастные группы.
Нельзя представить выраженность и направленность возрастных изменений любого типа гормональной регуляции только на основе знания концентрации гормона в крови или его секреции.
Необходим системный анализ, анализ с позиции саморегуляции, включающий характеристику всех звеньев системы, - гипоталамо-гипофизарной регуляции, активности железы, соотношение синтеза и распада гормонов, концентрации гормонов в крови, реакции эффекторов на действие гормона, обратных связей в системе гормональной регуляции. Концентрация гормонов в крови может изменяться при различных функциональных состояниях в широких пределах, и потому важно определение как чувствительности к гормонам (пороговые эффекторные концентрации) , так и реакционной способности (максимальный ответ на действие гормона). Системный анализ доказал неравномерные изменения в разных звеньях системы гормональной регуляции, связанные с одновременным развитием процессов старения и витаукта. Однако нарастающие с возрастом структурные и функциональные изменения, возникающие'во всех звеньях регулируемой системы, ведут в конечном итоге к ограничению ее надежности.
Появляются разнонаправленные изменения во всех звеньях системы тиреоидной регуляции (табл. 2). В старости наступают существенные изменения структуры и функции щитовидной железы, падает содержание в крови трийодтиронина и тироксина (Валуева, 1978). К старости нарушается способность специфических белков крови связывать тиреоидные гормоны (Фролькис и др., 1973). При введении гипоталамических рилизинг-факторов отмечено повышение чувствительности, но снижение реакционной способности подсистемы гипофиз-щитовидная железа у пожилых людей на действие гипоталамического фактора регуляции (Ohara et al., 1974). Предполагается, что нарушение транспорта тиреоидных гормонов связано с изменением состава плазмы, в частности преальбуминов, регулирующих влияние тироксина. Содержание ТТГ в аденогипофизе и крови снижается. Наряду с этим изменяется реактивность щитовидной железы к действию ТТГ. С возрастом растет чувствительность тиреоидной ткани к действию ТТГ, однако диапазон возможной ответной реакции щитовидной железы на введение ТТГ суживается, что ограничивает адаптационные возможности всей системы. Это установлено при изучении влияния различных доз ТТГ (от 5 до 35 ЕД/кг массы животного) на интенсивность потребления кислорода тканью железы и на содержание общего тироксина в крови (Фролькис и др., 1973; Вержиковская, Валуева, 1982).
У 18-24-месячных крыс высокая концентрация ТТГ в крови сохраняется при сниженной активности щитовидной железы. Это позволяет предположить, что возрастные изменения тиреотропной функции гипофиза не являются ведущими в процессе нарушения секреции щитовидной железы в старости. По данным нашей лаборатории, в старости изменяется реактивность тканей к действию тиреоидных гормонов. Так, оказалось, что в миокарде и печени старых крыс в более ранние сроки развития экспериментального гипертиреоза по сравнению со взрослыми повышается интенсивность потребления O2, разобщаются окисление и фосфорилирование, растет интенсивность гликолиза, падает содержание гликогена, изменяется содержание и обновление АТФ. Вместе с тем дача больших доз тиреоидных гормонов или их более длительное введение приводят к более выраженным изменениям у зрелых животных. По данным Г. В. Валуевой (1978), эти сдвиги в чувствительности и реакционной способности тканей определяются тем, что число рецепторов к тироксину в старости падает, а дейодирующая способность тканей растет. Например, у 28-32месячных крыс дейодирующая способность сердца и мышц почти вдвое больше, чем у 8-10-месячных животных.
Существенные изменения наступают и на этапе обратной связи - при введении малых доз тироксина более выраженное снижение содержания ТТГ наступает у старых крыс, а при больших - у зрелых. Итак, для системы тиреоиднои регуляции характерны неравномерные, порой разнонаправленные сдвиги в ее различных звеньях. Весь этот комплекс изменений способствует снижению тканевого дыхания, изменению сопряжения окисления и фосфорилирования, биосинтеза белка.
В процессе старения наступают существенные структурные нарушения в половых железах, прогрессивно снижается концентрация тестостерона у мужчин, а количество эстрадиола и прогестерона растет; у женщин падает содержание эстрадиола и прогестерона, а тестостерона растет. Введение хориогонина вызывало у старых людей более выраженный рост концентрации тестостерона, чем у пожилых. Одновременно снижается число рецепторов к половым стероидам, и это приводит к изменению реакции тканей. Возрастает также содержание ФСГ и ЛГ. По данным E. В. Мороз (1982), содержание ФСГ у женщин в 80-90 лет в 14 раз выше, а Л Г - в 5 раз выше, чем у 20-30-летних. Данные о содержании ФСГ, ЛГ и пролактина в крови мышей и крыс противоречивы. Важно, что в процессе старения резко изменяются количественные соотношения в содержании ФСГ и ЛГ и становится одной из причин нарушение репродуктивной функции. По мнению В. M. Дильмана (Дильман, Цырлин, 1982), рост содержания гонадотропных гормонов связан с повышением порога чувствительности гипоталамуса к действию половых стероидов.
На рис. 10 представлены данные E. В. Мороз и H. С. Верхратского (1984), характеризующие динамику изменения содержания половых стероидов и гонадотропных гормонов в крови мужчин разного возраста. Концентрация половых стероидов начинает снижаться раньше, чем нарастает содержание гонадотропных гормонов. Это позволяет полагать, что рост содержания гонадотропных гормонов - проявление витаукта, адаптивных реакций организма, связанных с усилением обратной положительной связи при снижении содержания тестостерона. Конечно, адаптивное значение этого сдвига ограничено и в определенных ситуациях он может приобретать патологическое значение. E. В. Мороз и H. С. Верхратский изучали реакцию гипоталамо-гипофизарной области на введение тестостерона в кровь. Оказалось, что малые дозы препарата подавляли секрецию гонадотропинов в более ранние сроки у пожилых людей, а большие дозы - более выраженные сдвиги у молодых.
Существенны изменения и на этапе прямой связи. По данным E. В. Мороз (1982), малые дозы хориогонина у старых животных более выраженно активируют функцию половых желез. Итак, в процессе старения наступают неравномерные изменения в разных звеньях саморегуляции функции половых желез, нарастающие возрастные изменения становятся причиной нарушения полового поведения, репродуктивной функции, биосинтеза белка и липидов в старости.

Рис. 10. Концентрация в крови ЛГ (/),ФСГ (2), эстрадиола (3), прогестерона тестостерона (5) у женщин (Л) и мужчин (Б) разного возраста.
В механизме метаболических нарушений при старении существенное значение имеет снижение инсулиновой обеспеченности организма, падение инсулиновой активности крови. Эти сдвиги способствуют развитию диабета в пожилом и старческом возрасте. Данные об изменении содержания инсулина в старости противоречивы. Одни исследователи наблюдают падение содержания инсулина в крови в старости, другие - его рост. У пожилых людей с нормальной толерантностью к глюкозе содержание инсулина практически не изменяется, а у людей с пониженной толерантностью- растет ((9.5HhI.9) у молодых, (14.8+1.8) мкЕД/мл у пожилых). Сочетание высокого уровня инсулина в крови у пожилых людей с низкой инсулиновой активностью может быть связано: 1) с увеличением содержания контраинсулярных факторов; 2) с появлением веществ, ингибирующих биологическую активность гормона; 3) с увеличением содержания проинсулина; 4) с накоплением антител к инсулину (Фролькис, 1977) В старости снижается содержание инсулина в ряде тканей и в эритроцитах (Никитин, 1982). В этих условиях адаптивное значение имеет снижение активности инсулиназы, изменение соотношения свободного и связанного гормона. В старости снижается возможный диапазон реакций тканей на действие инсулина, и это в значительной степени связано со снижением числа инсулинорецепторов. Вместе с тем реакция ряда тканей на малые дозы гормона длительно сохраняется. Специфический раздражитель для (3-клеток поджелудочной железы - сахар крови. По мнению одних исследователей, с возрастом снижается реакция р-клеток на гликемию, по мнению других, растет. По данным О. К. Кульчицкого и П. А. Орлова (1977), высокая гиперинсулинемия развивается у людей с низкой толерантностью к глюкозе. В старости ослабляется гипоталамический, нервный контроль над инсулярным аппаратом, и это снижает его адаптивные возможности. Снижение инсулиновой активности крови, затяжные гипергликемии, ингибирование биологической активности гормона способствуют развитию гиперинсулинемии, активации |3-клеток, что на фоне их структурных изменений ведет к развитию инсулиновой недостаточности, диабета у пожилых.
Цикл работ нашего коллектива об изменении инсулиновой обеспеченности организма в старости был обобщен в специальном сборнике «Инсулиновая обеспеченность организма в старости» (1977). Исходя из всех этих предпосылок, нами было предложено использование в терапии инсулиновой недостаточности веществ, разрывающих связь гормона с белком.
Существенное значение в ограничении адаптивных возможностей организма имеют сдвиги в системе гипоталамус-гипофиз- кора надпочечников-ткани. По нашим данным, концентрация альдостерона в крови в старости не изменяется, однако интенсивность секреции гормона падает (Магдич, 1983). Сохранение в этих условиях базального уровня гормона связано с ослаблением его распада и связывания белками.
Рис. 11. Секреция альдостерона и влияние на нее стимуляции латеральной гипоталамической области, гипофизэктомии и различных доз АКТГ а - контроль, б - стимуляция гипоталамуса, в - гипофизэктомия, г - АКТГ Светлые столбики и / - взрослые, заштрихованные и 2 - старые.
Как видно на рис 11, в старости ослабляется гипоталамо-гипофизарный контроль над секрецией альдостерона. Раздражение гипоталамуса, гипофизэктомия у старых крыс в меньшей степени влияют на секрецию альдостерона, однако чувствительность коры надпочечников к действию АКТГ и вазопрессина в старости растет, а реакционная способность падает. При старении падает число рецепторов, чувствительных к альдостерону, изменяется реакция тканей на действие гормона (Магдич, Верхратский, 1984).
Содержание кортикостерона, кортизола, по мнению одних исследователей, падает, других - не изменяется, увеличивается содержание свободной формы гормона (Свечникова и др , 1983). Концентрация АКТГ у людей не меняется, у крыс увеличивается. По нашим данным, в старости меньшие, пороговые дозы АКТГ активируют кору надпочечников, однако при больших дозах стимуляция более выражена у зрелых животных. Подобные изменения реактивности характеризуют реакцию тканей на действие кортикостерона. Существенные изменения происходят и на этапе обратной связи: при введении 100 мкг дексазон i на 1 кг массы ст,виг в содержании АКТГ более выражен \ зрелых крыс, а при введении 500 мкг/кг - у старых
Существенно изменяется синтез и обмен вазопрессина. Цикл наших работ был обобщен в специальной книге «Вазопрессин и патология сердечно-сосудистой системы в старости» (1983). Оказалось, что у пожилых людей, старых животных растет концентрация гормона в крови, определенная как биологическими, гак и радиоиммунологическими методами (25-летние мужчины - (24±0.3); 65-75-летние- (102±18) пмоль/мл). Этот рост концентрации связан как с усилением синтеза вазопрессина, так и снижением интенсивности его распада. Наряду с этим растет чувствительность сердца и сосудов к действию гормона. При введении вазопрессина в дозе 0.005 E Д/100 г у зрелых крыс не возникает сдвигов ЭКГ, а у старых в 65 % случаев отмечается увеличение зубца T, в 40 % - смещение вверх сегмента S-T. У старых собак вазопрессин в дозах, в 2 раза меньших, чем у взрослых животных, вызывал спазм коронарных сосудов. Кроме того, в старости ослабляется тахифилаксия на вазопрессин (Медведь, 1980). Эта триада - рост концентрации гормона, повышение чувствительности к нему сердечно-сосудистой системы, ослабление тахифилаксии - способствует развитию возрастной патологии - артериальной гипертонии, коронарной недостаточности (Фролькис и др., 1984). И, действительно, по данным С. Ф. Головченко (1983), у больных артериальной гипертонией, ишемической болезнью сердца нарастает концентрация гормона в крови. Исходя из этих предпосылок, нами и было предложено использовать ингибиторы вазопрессина в терапии этих заболеваний.
Итак, в любой системе гормональной регуляции возникают разнонаправленные сдвиги, зачастую компенсирующие друг друга, - сочетание снижения концентрации гормона с ростом чувствительности к нему, изменение соотношения свободных и связанных форм гормонов, снижение интенсивности секреции и ослабление процесса распада гормона, активация ряда гипоталамических механизмов и др. Адаптивное значение имеет определенное соотношение между метаболическими возможностями тканей и уровнем их гормональной регуляции в старости, ибо чрезвычайная гормональная активация многих метаболически затухающих процессов привела бы к катастрофическим последствиям. Вместе с тем изменения в механизмах секреции, в реактивности тканей и самих желез, изменение обмена гормонов приводят к нарастающей недостаточности ряда гормональных систем. Одним из важнейших механизмов в этих сдвигах является изменение рецепции клеток во всех звеньях системы. Своеобразная «дерецепция» клеток в старости изменяет межклеточные отношения, снижает базальный уровень обмена. Одна и та же клетка находится под контролем различных гормонов. При старении неодинаково изменяется число рецепторов в клетке к различным гормонам, и это становится важной причиной изменения реактивности клеток в старости (Roth, 1979). Сопоставление возрастных изменений концентрации гормонов в крови и рецепции к ним клеток доказывает, что гормональный контроль в старости существенно изменяется не только количественно, но и качественно. Изменяется участие различных гормонов в регуляции клеточного метаболизма. Это изменение реактивности клеток определяется не только сдвигами в количестве рецепторов, но и в их свойствах, связанных во многом с конформационными изменениями молекул, со сдвигами в липидном окружении и др.
С возрастом изменяются не только отдельные системы гормональной регуляции, но и межэндокринные связи в единой системе нейрогормонального контроля. Изменяется влияние катехоламинов на гипоталамус и гипофиз, половых стероидов на функцию коры надпочечников, тиреоидных гормонов на обмен катехоламинов и др. Важное значение в межэндокринных связях имеет феномен гормонального подавления. В нашем коллективе было показано, что увеличение концентрации ряда тропных гормонов в старости подавляет секрецию соответствующих желез. Так, большое количество гонадотропных гормонов у старых крыс подавляет секрецию тестостерона, а кортикотропина - секрецию альдостерона и кортикостерона. Иными словами, активирующие гормональные влияния могут стать угнетающими, что может быть одним из механизмов снижения секреторной активности желез внутренней секреции. Это положение приемлемо и при оценке влияния гормонов на органы. Вазопрессин, катехоламины в дозах, вызывающих оптимальный сдвиг функции сердечно-сосудистой системы у взрослых крыс, приводят к некрозам у старых. Гидрокортизон в дозах, вызывающих оптимальную индукцию ряда ферментов печени у взрослых животных, угнетает ее у старых. Гормональные регуляторные влияния могут в старости вызывать повреждающие эффекты - возникает рассогласование между регуляторным сигналом, идущим по прямой связи, и реактивностью объекта регуляции.
См. также:
 Интегративная деятельность мозга
Интегративная деятельность мозга
Лимбическая система и гипоталамус
Нейротрофические механизмы
Обратные связи в системе нейрогуморальной регуляции
Изменения реактивности эффекторов
 Обсудить на форуме
Обсудить на форуме

